Lipidenvertering en -absorptie zijn complexe processen. Er zijn oplosbare enzymen bij betrokken, substraten met verschillende oplosbaarheidsgraden, en ze vinden voornamelijk plaats in de maag en de dunne darm.
Lipiden in de voeding zijn triglyceriden, fosfolipiden, steroïden, vooral cholesterol en cholesterolesters, vetoplosbare vitaminen, namelijk vitamine A, D, E en K, en carotenoïden.
Lipiden kunnen bij kamertemperatuur vast of vloeibaar zijn en worden respectievelijk vetten en oliën genoemd.
- Triglyceriden of triacylglycerolen maken ongeveer 90% van de voedingslipiden uit. Zij bestaan uit één glycerolmolecuul veresterd met drie vetzuren, meestal vetzuren met een lange keten (16-20 koolstofatomen). Zij hebben een energiedichtheid die meer dan twee keer zo groot is als die van koolhydraten (9 kcal/g tegenover ongeveer 3,75) en aminozuren. Ze moeten hun vetzuren afgeven om als energiebron te worden gebruikt.
- Fosfolipiden, de belangrijkste bestanddelen van biologische membranen, bestaan uit één glycerolmolecuul veresterd met twee vetzuren op de sn-1 en sn-2 posities, en een fosforzuur op de sn-3 positie. De fosfaatgroep bindt op zijn beurt een hydrofiele groep, zoals choline, serine of inositol, via een esterbinding. De dagelijkse inname van fosfolipiden is laag, 1-2 g; er komen echter ook biliaire fosfolipiden in de dunne darm terecht, ongeveer 10-20 g per dag, voornamelijk fosfatidylcholine.
- Cholesterol en zijn esters komen, samen met kleine hoeveelheden steroïdhormonen, alleen voor in dierlijke produkten, in tegenstelling tot de tot nu toe geziene lipiden die ook in plantaardige produkten voorkomen.
In de dunne darm komt, naast cholesterol uit de voeding (die niet meer dan 300 mg/dag mag bedragen), ook galcholesterol, ongeveer 1 g/dag. Zowel cholesterol in de voeding als cholesterol in de gal zijn meestal in niet-veresterde vorm, ongeveer 85-90%, de enige vorm van cholesterol die in de dunne darm kan worden opgenomen.
Net als vitamines is ook dit steroïde molecuul geen bron van energie.
Een verscheidenheid aan plantenstanolen en -sterolen, met name het β-sitosterol (dat onder fysiologische omstandigheden niet wordt geabsorbeerd), wordt ook tot de voedingssteroïden gerekend.
Ondanks dat wetenschappelijke genootschappen een vetinname (in principe triacylglycerolen) van niet meer dan 30% van de dagelijkse calorie-inname aanbevelen, leveren in de westerse voeding vetten en oliën tussen de 30 en 45% van de dagelijkse calorie-inname.
Hydrofobiciteit, een van de onderscheidende eigenschappen van veel voedingslipiden, die triglyceriden tot uitstekende moleculen voor energieopslag maakt, levert problemen op wanneer dergelijke moleculen in het maagdarmkanaal worden verteerd, in de dunne darm worden geabsorbeerd, en tenslotte in de circulatie worden getransporteerd na absorptie of mobilisatie uit lichaamsvoorraden.
Vetten zoals triglyceriden met lange keten vetzuren, en cholesterol en vetoplosbare vitamine-esters zijn namelijk extreem hydrofoob, en aggregeren tot grote druppels in de maag en de dunne darm. Deze druppels worden dan geëmulgeerd om hydrolasen in staat te stellen de vertering van lipiden te katalyseren.
CONTENTEN
- Stappen, enzymen en producten
- Lipidenvertering en linguale lipase
- Lipidenvertering en maaglipase
- Galzouten en de emulsie van lipidedruppels
- Lipidenvertering en pancreaslipase
- Lipidenvertering en lipase in moedermelk
- Lipidenvertering en cholesterolesterase
- Lipidenvertering en fosfolipasen
Stappen, enzymen, en producten
De vertering van lipiden begint in de mond, gaat verder in de maag, en eindigt in de dunne darm.
Enzymen die betrokken zijn bij de vertering van triacylglycerol worden lipase (EC 3.1.1.3) genoemd. Het zijn eiwitten die de gedeeltelijke hydrolyse van triglyceriden tot een mengsel van vrije vetzuren en acylglycerolen katalyseren. Er zijn verschillende lipasen, waarvan de belangrijkste wordt geproduceerd door de exocriene pancreas; de andere zijn linguale lipase, maaglipase en moedermelklipase.

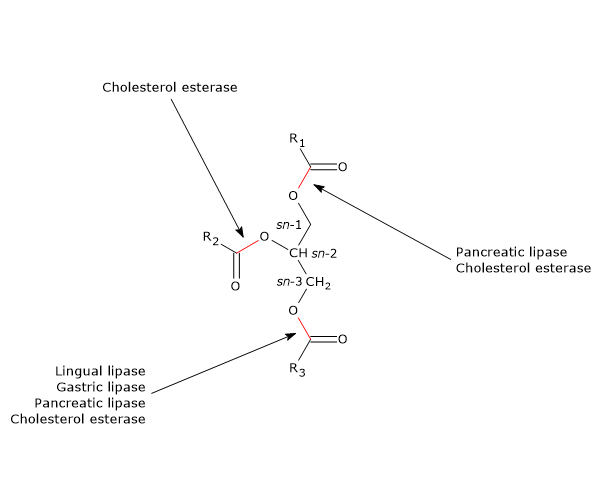
Andere enzymen die betrokken zijn bij de vertering van lipiden zijn cholesterolesterase en fosfolipasen A1 en A2.
Lipidenvertering en linguale lipase
In de mond wordt voedsel in kleine deeltjes gebroken en vermengd met linguale lipase.
Het enzym wordt geproduceerd en uitgescheiden door sereuze linguale klieren, ook wel von Ebner’s klieren genoemd.
Het is stabiel in een zure omgeving en blijft daarom actief in de maag, en ook in de dunne darm in het geval dat er geen goede pancreas afscheiding van bicarbonaat is.
Bij de reactie die door het enzym wordt gekatalyseerd, komt één vetzuur vrij, bij voorkeur een korte- of middellange-keten vetzuur, en een 1,2-diacylglycerol, dat vervolgens in de twaalfvingerige darm wordt gehydrolyseerd.
Notitie: korte-keten vetzuren zijn voornamelijk veresterd in de sn-3 positie van het triacylglycerol.
Linguale lipase speelt een bescheiden rol in de vertering van triacylglycerolen, omdat:
- het een langzamere activiteit heeft dan pancreaslipase;
- het werkt in de maag, een waterige omgeving waarin de lipiden de neiging hebben samen te smelten, en een aparte fase te vormen van de omgeving, waardoor de mogelijkheden voor het enzym om triacylglycerolen te hydrolyseren beperkt worden.
Linguale lipase-activiteit is niet bijzonder belangrijk voor gezonde volwassenen. Integendeel, zij is zeer belangrijk voor zuigelingen, bij wie de pancreaslipase nog onvolgroeid is, mede bevoordeeld door het feit dat de triglyceriden in melk rijk zijn aan vetzuren met korte en middellange keten. Bovendien kan het, net als maaglipase (zie hieronder), doordringen tot in de vetbolletjes van de melk, en zo het verteringsproces op gang brengen (pancreaslipase kan niet doordringen tot in deze vetbolletjes).
Het is duidelijk dat de werking van dit enzym ook belangrijk is wanneer pancreaslipase afwezig is.
Aangezien de tong gevoelig is voor de smaak van vrije vetzuren, vooral meervoudig onverzadigde, in plaats van triglyceriden, zou linguale lipase activiteit een rol kunnen spelen bij de detectie van vet voedsel als energiebron, en daarmee de voedselkeuze kunnen beïnvloeden.
Ten slotte is het vrijkomen van korte- en middellange-keten vetzuren en diacylglycerolen ook van belang omdat het amfipathische moleculen zijn, dat wil zeggen dat ze een hydrofiel gedeelte hebben, dat in wisselwerking staat met de omringende waterige fase, en een hydrofoob gedeelte, dat naar de kern van de lipidedruppels is georiënteerd. Door de werking van deze oppervlakte-actieve stoffen krijgen de vetdruppels een hydrofiel oppervlak, d.w.z. een stabiel raakvlak met de omringende waterige fase. Dit leidt, samen met de kolkende werking van de maag, tot de vorming van een emulsie van druppeltjes, die in omvang afnemen. De emulsie komt vervolgens als chyme in het duodenum terecht. Andere amfipathische moleculen die in voedsel aanwezig zijn, zijn lecithine en fosfolipiden, die samen het oppervlak vergroten dat beschikbaar is voor de hydrolase-activiteit.
In de maag worden de lipiden ook vermengd met maagsap en zoutzuur. Het zuur denatureert ook de eiwitten van de lipide-eiwit complexen; de polypeptiden worden dan verteerd door maagproteasen, en de lipiden komen vrij.
Lipide vertering en maaglipase
In de maag worden de lipiden ook gehydrolyseerd door een tweede zuur lipase, het maaglipase. Dit enzym wordt uitgescheiden door de hoofdcellen van het maagslijmvlies, en heeft een optimale pH rond 4, maar is nog vrij actief bij minder zure pH-waarden, 6 tot 6,5. Daarom blijft het waarschijnlijk ook actief in het bovenste deel van de twaalfvingerige darm, waar de pH tussen 6 en 7 ligt.
Het enzym katalyseert bij voorkeur de hydrolyse van triglyceriden met korte- en middellange-keten vetzuren, maar kan ook lange-keten vetzuren hydrolyseren. Ongeacht het type vetzuren, splitst maaglipase bij voorkeur die op de sn-3 positie, wat leidt tot het vrijkomen van een vrij vetzuur en een 1,2-diacylglycerol, moleculen die kunnen fungeren als oppervlakte-actieve stoffen, zoals eerder is gebleken.
Zoals linguale lipase is het vooral actief op triglyceriden van melk, ook van moedermelk, die rijk zijn aan vetzuren met korte en middellange keten. Bovendien is het ook actief op triacylglycerolen van sommige tropische oliën, bijvoorbeeld kokosolie, die rijk zijn aan middellange-keten vetzuren.
Het enzym kan 10 tot 30% van de triacylglycerolhydrolyse in het maagdarmkanaal voor zijn rekening nemen, en tot 50% bij zuigelingen die borstvoeding krijgen.
De galzouten en de emulsie van lipidedruppels
Het chyme, dat een lipide-emulsie bevat die bestaat uit druppels met een diameter van minder dan 0,5 mm, komt in het bovenste deel van de dunne darm, de twaalfvingerige darm, waar de hydrolyse van triglyceriden wordt voortgezet.

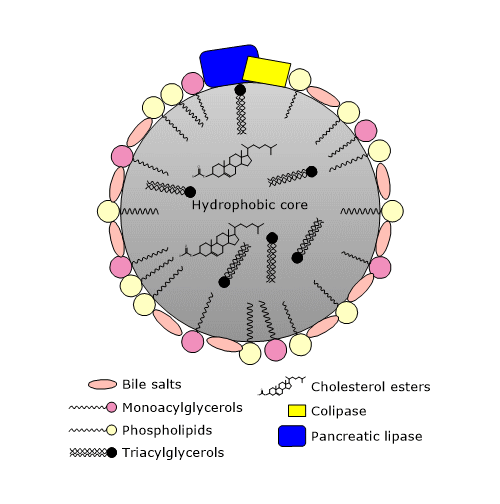
In de twaalfvingerige darm, wordt het chym vermengd met gal, waarvan de afgifte door de galblaas wordt gestimuleerd door cholecystokinine, een hormoon dat wordt afgescheiden door cellen van het slijmvlies van de twaalfvingerige darm en het jejunum als reactie op de inname van een maaltijd, vooral als die veel vet bevat. In de gal bevinden zich onder andere galzouten, fosfolipiden en cholesterol. Galzouten zijn galzuren die geconjugeerd zijn met glycine of taurine. Galzuren zijn op hun beurt zuurstofhoudende derivaten van cholesterol. Galzuren en galzouten worden beide door de lever gesynthetiseerd. Het zijn amfipathische moleculen, in wier planaire ringstructuur een hydrofoob en een hydrofiel vlak te herkennen zijn. Daarom zijn ze in staat lipidedruppels verder te emulgeren, waardoor het oppervlak voor hydrolaseactiviteit wordt vergroot.
Met name zouten van cholzuur, die drie hydroxylgroepen bevatten, zijn betere emulgatoren dan zouten van deoxycholzuur, die in plaats daarvan slechts twee hydroxylgroepen bevatten.
Notitie: de galblaas scheidt elke dag ongeveer 30 g galzouten af, samen met fosfolipiden en cholesterol. Het grootste deel van de galzouten en cholesterol wordt vervolgens weer geabsorbeerd, zodat het dagelijkse fecale verlies van galzouten en steroïden vrij laag is, 0,2-1 g.
Het mechanisme van peristaltiek en de tot nu toe geziene oppervlakteactieve stoffen (vrije vetzuren, acylglycerolen, fosfolipiden, en galzouten) zorgen voor de vorming van microscopisch kleine micellen, die het beschikbare oppervlak voor hydrolytische enzymactiviteiten nog vergroten.
Het moet worden benadrukt dat triacylglycerolen met korte- en middellange-keten vetzuren zowel kunnen worden gehydrolyseerd als geabsorbeerd in afwezigheid van galzouten, hoewel hun aanwezigheid de absorptie verhoogt.
Lipidenvertering en pancreaslipase
Cholecystokinine stimuleert ook de exocriene pancreas tot de afscheiding van een pancreassap dat, naast andere moleculen, pancreaslipase bevat. Dit enzym katalyseert de vertering van het merendeel van de ingenomen triglyceriden, voornamelijk in het bovenste deel van het jejunum, en heeft een optimale pH van 7,0 tot 8,8 (het is dus geen zure lipase, zoals linguale en maaglipases). Het katalyseert de splitsing van vetzuren, gewoonlijk met 10 of meer koolstofatomen, voornamelijk in de sn-1 en sn-3 posities van de glycerolruggengraat. De producten van de reactie zijn vrije vetzuren en 2-monoacylglycerolen. Het 2-monoacylglycerol, de belangrijkste vorm waarin de monoacylglycerolen uit de dunne darm worden opgenomen, kan een isomerisatieproces ondergaan waarbij het resterende vetzuur naar koolstof 1 of 3 verhuist. De snelheid van de isomerisatie is echter trager dan de snelheid van de opname van de molecule uit de dunne darm. In vitro wordt pancreaslipase geremd door galzouten, terwijl het in vivo triglyceriden op zeer efficiënte wijze hydrolyseert, dankzij de aanwezigheid van een eiwit-cofactor die door de exocriene pancreas wordt afgescheiden, het colipase. Dit eiwit heeft geen katalytische activiteit, wordt geproduceerd in inactieve vorm, procolipase genaamd, en wordt geactiveerd door trypsine in de twaalfvingerige darm. Lipidedruppels zijn gecoat met fosfolipiden en galzouten, die hen een negatieve lading geven die de binding van lipase verhindert, maar colipase aantrekt. Colipase bindt op zijn beurt pancreaslipase (lipase en colipase binden in een molaire verhouding van 1:1), waardoor het enzym wordt verankerd aan het water-lipide grensvlak van de lipidedruppels.
Lipide-vertering en lipase in moedermelk
Dit lipase heeft een neutraal pH-optimum, en wordt gestimuleerd door galzouten. Het enzym draagt in belangrijke mate bij tot de hydrolyse van de triglyceriden in de darm van zuigelingen die borstvoeding krijgen.
In koemelk komen daarentegen geen lipasen voor.
Lipidenvertering en cholesterolesterase
Een ander enzym dat in het pancreassap aanwezig is, is cholesterolesterase (EC 3.1.1.13). Dit enzym, dat door de exocriene pancreas wordt gesynthetiseerd en in actieve vorm wordt uitgescheiden, is een lipase met een brede specificiteit, die actief is op:
- cholesterolesters, met cholesterol en vrije vetzuren als reactieproducten;
- triglyceriden, waarin het alle drie de esterbindingen hydrolyseert, en om deze reden wordt het ook niet-specifiek esterase genoemd (het hydrolyseert 10%-15% van de triglyceriden in de voeding);
- monoacylglycerolen;
- fosfolipiden;
- esters van vitamine A en D.

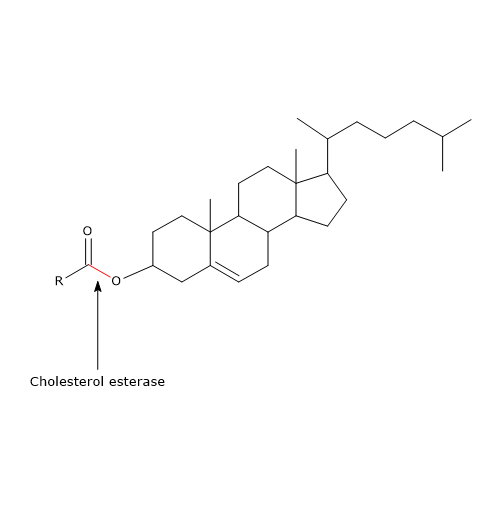
Zoals fosfolipase A2 (zie hieronder) is het vooral actief op cholesterolesters die in galzoutmicellen zijn opgenomen. In tegenstelling tot pancreaslipase wordt zijn activiteit gestimuleerd door galzouten, voornamelijk trihydroxyzouten, zoals natriumtaurocholaat en glycocholaat. Deze zouten veroorzaken een verandering in de conformatie van het eiwit, waardoor het enzym wordt geactiveerd. Bovendien bevorderen trihydroxyzouten de zelf-associatie van het enzym tot polymere aggregaten, die het beschermen tegen de werking van proteasen in het darmlumen.
Lipidenvertering en fosfolipasen
De vertering van fosfolipiden wordt uitgevoerd door fosfolipasen, voornamelijk fosfolipase A2 (EC 3.1.1.4). Het enzym is aanwezig in het pancreassap in de vorm van een zymogeen, prophospholipases A2 genaamd, en wordt geactiveerd door trypsine , een enzym dat betrokken is bij de vertering van eiwitten. Fosfolipase A2 katalyseert specifiek de splitsing van het vetzuur op de sn-2 positie van de fosfolipiden, terwijl het een brede specificiteit heeft met betrekking tot zowel de lengte van de koolstofketen van het doel-vetzuur als de polaire kopgroepen van de fosfolipiden.
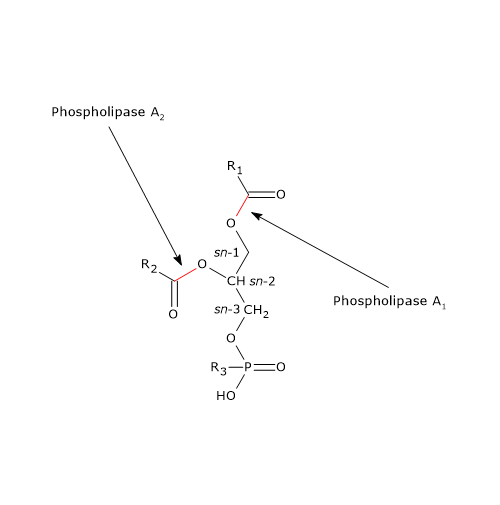
Zoals eerder gezien, zijn de meeste fosfolipiden in het darmlumen afkomstig van de gal en slechts een klein deel van de voeding. In de gal vormen de fosfolipiden micellen met cholesterol en galzouten, en in het darmlumen worden ze verdeeld tussen de lipidedruppels en deze micellen, met een voorkeur voor de laatste. En in de micellen fungeren fosfolipiden, meestal fosfatidylcholine, als substraat. In het geval van fosfatidylcholine zijn een vrij vetzuur en lysofosfatidylcholine (een lysofosfolipide) de reactieproducten.
In pancreassap is ook fosfolipase A1 aanwezig, dat het vetzuur op de sn-1 positie van de fosfolipide verwijdert.
In het darmslijmvlies lijkt er nog een derde, bescheiden, fosfolipase activiteit te zijn, dankzij een intrinsiek membraaneenzym. Dit enzym wordt fosfolipase B of retinyl ester hydrolase genoemd, en is ook actief op vitamine A esters.
De vertering van fosfolipiden kan eindigen met de vorming van een vrij vetzuur en een lysofosfolipide of kan volledig zijn.
Bender D.A. Introduction to nutrition and metabolism. 3e Editie. Taylor & Francis, 2004
Berdanier C.D., Dwyer J., Feldman E.B. Handbook of nutrition and food. Tweede editie. CRC Press. Taylor & Francis Group, 2007
Chow Ching K. “Fatty acids in foods and their health implication” 3e Editie. CRC Press Taylor & Francis Group, 2008
Hamosh M. Lingual and gastric lipases. Nutrition 1990;6(6):421-8.
Iqbal J. and Hussain M.M. Intestinal lipid absorption. Am J Physiol Endocrinol Metab 2009;296:E1183-94. doi:10.1152/ajpendo.90899.2008
Nelson D.L., Cox M.M. Lehninger. Beginselen van de biochemie. 4e Editie. W.H. Freeman and Company, 2004
Rosenthal M.D., Glew R.H. Medical Biochemistry – Human Metabolism in Health and Disease. John Wiley J. & Sons, Inc., Publicatie, 2009
Stipanuk M.H., Caudill M.A. Biochemical, physiological, and molecular aspects of human nutrition. 3e Editie. Elsevier gezondheidswetenschappen, 2013