Inleiding
Neuronen ontvangen voortdurend van duizenden andere neuronen in hun omgeving. Of deze inputs al dan niet een actiepotentiaal kunnen opwekken, hangt echter af van de sommatie van deze inputs. Sommering kan worden gedefinieerd als een proces waarbij de excitatoire en inhibitoire signalen samen in staat zijn al dan niet een actiepotentiaal op te wekken.

Er zijn twee soorten sommaties, dit zijn respectievelijk de temporele en de ruimtelijke sommaties. Ruimtelijke sommaties kunnen worden gedefinieerd als signalen die afkomstig zijn van meerdere gelijktijdige ingangen. Temporele sommatie daarentegen is afkomstig van herhaalde inputs. Om een actiepotentiaal te bereiken, moet in wezen de drempelspanning worden bereikt. Deze kan worden bepaald door de afzonderlijke inputs van temporele en ruimtelijke sommatie bij elkaar op te tellen.
Er zijn twee soorten neurotransmitters die vrijkomen uit presynaptische neuronen. Depolarisatie wordt veroorzaakt door excitatoire neurotransmitters, dit wordt een excitatoire postsynaptische potentiaal (EPSP) genoemd. Inhibitoire neurotransmitters veroorzaken hyperpolarisatie, met andere woorden een inhibitory postsynaptic potential (IPSP). Neuronen kunnen elkaar op een aantal manieren beïnvloeden. Zij kunnen excitatie en inhibitie veroorzaken en kunnen elkaars exciteerbaarheid beïnvloeden.
Defacilitering is het wegnemen van exciterende inputs die een pad kunnen faciliteren. Disinhibitie daarentegen is het verwijderen van remmende inputs.
Meervoudige broningangen op een neuron kunnen ruimtelijk worden samengevat, maar de ingangen moeten dicht bij elkaar liggen, zodat geen van de vroege ingangen vervalt. Als een neuron meerdere ingangen van één bron ontvangt in het vereiste nauwe tijdsinterval, zodat geen enkele ingang vervalt, dan kunnen deze ingangen temporeel worden opgeteld.
Een andere zeer belangrijke factor bij het bepalen of de drempelpotentiaal wordt bereikt en een actiepotentiaal wordt bereikt of niet, is de afstand tussen de synaps en het neuronale cellichaam. Hoe dichter de afstand tussen de synaps en het cellichaam, des te groter is de sommatie en des te waarschijnlijker is het dat een actiepotentiaal wordt opgewekt.
Zoals we weten is het vervoermiddel voor een postsynaptisch neuron een dendriet. Deze dendrieten hebben weinig spanningsafhankelijke ionkanalen. Vandaar dat bij het bereiken van het neuronale cellichaam de postsynaptische potentiaal verzwakt. Dit neuronale cellichaam vat deze inkomende potentialen samen. Een actiepotentiaal wordt dan opgewekt bij transmissie van de nettopotentiaal naar de axonheuvel.
Spatiale sommatie
Wanneer inputs van meerdere neuronen een actiepotentiaal opwekken, wordt dit ruimtelijke sommatie genoemd. Deze potentialen zijn meestal afkomstig van dendrieten, we tellen deze ingangen bij elkaar op om de ruimtelijke sommatie te krijgen.
Hoe groter het aantal excitatoire post-synaptische potentialen, des te groter de kans dat de potentiaal de drempelwaarde bereikt om een actiepotentiaal op te wekken. Evenzo, hoe groter het aantal remmende postsynaptische potentialen, des te kleiner de kans dat de potentiaal de drempelpotentiaal bereikt om een actiepotentiaal op te wekken.
De kans dat een actiepotentiaal wordt opgewekt, wordt ook aanzienlijk beïnvloed door hoe dicht de dendritische ingang zich bij de axonheuvel bevindt. Hoe dichter de dendritische ingang zich bij de axonheuvel bevindt, des te waarschijnlijker is het dat deze een actiepotentiaal opwekt. Shunting van een excitatoir postsynaptisch potentieel is de tenietdoening van een excitatoire input door de ruimtelijke sommatie van remmende inputs.
Temporele sommatie
Wanneer een grote hoeveelheid presynaptische neuron-actiepotentialen postsynaptische actiepotentialen teweegbrengt die met elkaar sommeren, wordt dit temporele sommatie genoemd.
In dit geval is het interval tussen de actiepotentialen korter dan de postsynaptische actiepotentiaalduur. De sommatie kan worden vergroot als de tijdconstante van het celmembraan lang genoeg is. Wanneer de volgende postsynaptische potentiaal begint, zal de amplitude van de vorige postsynaptische potentiaal ermee worden samengeteld, waardoor een grotere potentiaal ontstaat die de kans vergroot dat de drempelpotentiaal wordt bereikt.
Mechanisme
De postsynaptische cellen bevatten ionkanalen, deze ionkanalen kunnen zich openen of sluiten, afhankelijk van welke neurotransmitter zich aan de receptoren bindt. Het openen/sluiten van deze kanalen creëert een postsynaptische potentiaal. Er zijn 2 soorten postsynaptische potentialen. Een exciterende postsynaptische potentiaal is die welke de kans op het initiëren van een actiepotentiaal verhoogt. Een remmende postsynaptische potentiaal verkleint de kans op het ontstaan van een actiepotentiaal.
Excitatoire neurotransmitters (Glutamaat)
Een uitstekend voorbeeld van een excitatoire neurotransmitter is glutamaat. Dit glutamaat bindt zich aan AMPA-receptoren op het postsynaptische membraan. Deze binding veroorzaakt een instroom van natriumkationen. Deze natrium influx veroorzaakt een depolarisatie. Dit wordt de excitatoire postsynaptische potentiaal (EPSP) genoemd. Het is van essentieel belang op te merken dat voor de EPSP-sommatie om de drempelpotentiaal te bereiken, een groot aantal van deze inputs nodig is. De effecten van neurotransmitters duren veel langer dan die van presynaptische impulsen.
Het verschil tussen excitatoire postsynaptische potentialen en actiepotentialen is dat excitatoire postsynaptische potentialen in staat zijn hun inputs samen te vatten, waardoor een trapsgewijze respons ontstaat, in tegenstelling tot de alles of niets respons waarbij de drempelpotentiaal kan worden bereikt door het stimuleren van een actiepotentiaal of helemaal niet wordt bereikt.
Remmende neurotransmitters (GABA)
GABA is de belangrijkste neurotransmitter die betrokken is bij remmende postsynaptische potentialen (IPSP). Bij binding aan postsynaptische neuronreceptoren opent GABA specifieke ionenkanalen, die verschillen van de kanalen die bij EPSP worden geopend door excitatoire neurotransmitters zoals glutamaat. Deze kanalen maken een instroom van negatief geladen anionen of een uitstroom van positief geladen kationen mogelijk. Het anion in dit geval zijn chloride-ionen. De uitgescheiden kationen zijn kalium-ionen. Beide ionen hebben hetzelfde effect in het verlagen van de membraanpotentiaal waardoor hyperpolarisatie van het postsynaptische neuron optreedt.
Sommatie van deze IPSPs en de daling van de membraanspanning zal afwijken van de drempelpotentiaal waardoor een actiepotentiaal wordt geremd. Deze IPSPs en EPSPs kunnen echter tegelijkertijd optreden, zodat het postsynaptische neuron excitatoire signalen van glutamaat en inhibitoire signalen van GABA kan ontvangen. Het doel van remmend glutamaat is om de membraanpotentiaal weg van de drempelpotentiaal te verlagen door hyperpolarisatie.
Algebraïsche verwerking van EPSP’s en IPSP’s
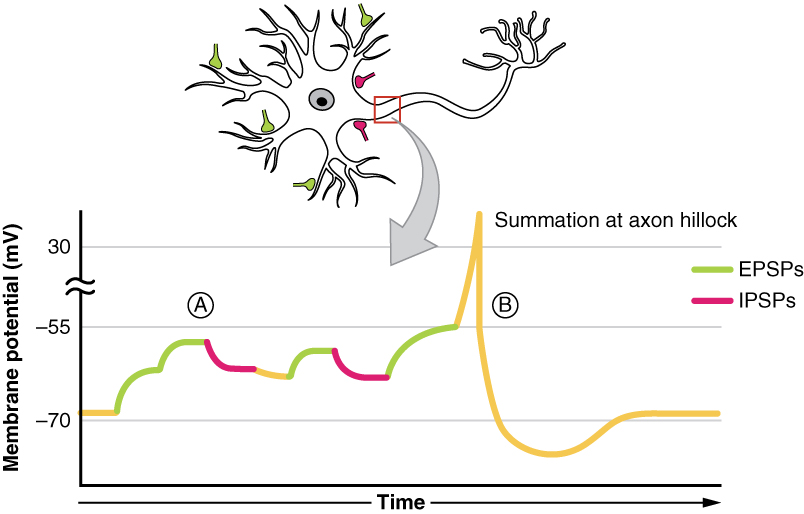
In een neuron zullen op elk willekeurig moment talrijke EPSP- en IPSP-inputs tegelijk worden ontvangen. Om de uitvoer te bepalen of de drempelpotentiaal zal worden bereikt en een actiepotentiaal zal worden opgewekt of niet, moet de algebraïsche verwerking van deze EPSP’s en IPSP’s worden genomen. Deze neuronen ontvangen talrijke inputs, hetzij van meerdere neuronen (ruimtelijke sommatie), hetzij van één enkel neuron (temporele sommatie).
Deze output hangt af van het aantal van elk type neurotransmitter, of het nu gaat om excitatoire neurotransmitters zoals glutamaat, die een instroom van natriumionen veroorzaken via natriumionkanalen, of remmende neurotransmitters zoals GABA, die een instroom van chloride-ionen of kalium-ionen veroorzaken via respectievelijk chloride- of kaliumionkanalen. Deze synaps kan worden aangeduid als het beslissingspunt waar de algebraïsche verwerking van deze IPSPs en EPSPs de output bepaalt.
Axonheuvel

Het deel van het cellichaam van een neuron dat in verbinding staat met het axon, wordt de axonheuvel genoemd. Het heeft een spaarzame verdeling van Nissl-substantie. We kunnen het met lichtmicroscopie identificeren. Aangezien deze axonheuvel de axon en de soma van het neuron verbindt, is het de laatste regio van de soma waar de sommering van membraanpotentialen van synaptische inputs plaatsvindt. Deze sommatie wordt vervolgens doorgegeven aan het axon.
Hoewel in het verleden velen geloofden dat de axonheuvel de trigger-zone was waar actiepotentialen worden geïnitieerd, gelooft men nu dat het initiële segment tussen het eerste ongemyeliniseerde axonsegment en de axonheuvelpiek de plaats is waar de actiepotentiaal wordt geïnitieerd. Het positieve punt van het axon waar de actiepotentiaal wordt gestart varieert van cel tot cel.
Hormonale stimulatie en neurotransmitter second messenger effecten kunnen dit positieve punt ook veranderen. Lokalisatie van membraanpotentialen naar het somale of axonale deel van de cel kan worden bereikt door afbakening van verschillende membraandomeinen tussen het axon en het cellichaam. Dit kan worden bereikt door de axonheuvel.
Shunting
Vaak liggen in een neuron de excitatoire postsynaptische en inhibitoire postsynaptische potentialen in een dendriet zeer dicht bij elkaar. Dit wordt shunting genoemd. Behalve in dendrieten kan ook in het soma van een cel shunting optreden.
Temporele sommatie zou ons vertellen dat we deze excitatoire postsynaptische en inhibitoire postsynaptische potentialen moeten sommeren om de resulterende output te bepalen of de drempelpotentiaal wordt bereikt en een actiepotentiaal wordt geïnitieerd of niet. Wanneer deze opeenvolging van gebeurtenissen zich echter in het soma van een roep voordoet, wordt de celweerstand veranderd door de remmende input. De cel begint te lekken, waardoor een shunt ontstaat in tegenstelling tot het wegnemen van het effect van de excitatoire input.
Therapeutische toepassing
Wanneer we het hebben over nociceptieve stimulatie, is temporele sommatie de repetitieve integratie van pijnprikkels. Ruimtelijke sommatie daarentegen is nociceptieve input integratie van grote gebieden. Veel chronische ziekten hebben symptomen van langdurige pijn die wijdverspreid is. Vandaar dat zowel temporele als ruimtelijke nociceptieve sommaties worden aangetroffen bij chronische ziekten. Experimenten met drukstimulatie leveren het bewijs dat de temporele sommatie van nociceptieve inputs wordt vergemakkelijkt door ruimtelijke sommatie. Daarom is de beste manier om chronische pijn te behandelen het richten van de behandeling op zowel ruimtelijke als temporele sommatie van pijn.
Onderzoek
De meeste experimenten van ruimtelijke sommatie worden getest op optische en sensorische neuronen, de reden hiervoor is dat zij een constant frequentiebereik hebben van excitatoire en inhibitoire neuronen. Postsynaptische potentiaal verzwakking van neuron cellichamen en dendrieten is grote focus van neurale sommatie in recente dag studies. Wegens het feit dat de respons van deze interacties minder is dan de som van de individuele responsen, worden deze responsen als niet-lineair beschreven. Shunting is een veel voorkomende oorzaak van dit effect. Shunting is een verminderde excitatoire postsynaptische potentiaalgeleiding.
Naoki Kogo en Michael Ariel waren in staat om experimenten uit te voeren op de basale optische kern van schildpadden, waaruit zij informatie konden putten over shunting inhibitie. Volgens hun werk veroorzaakte EPSP en IPSP ruimtelijke sommatie excitatoire responsverzwakking tijdens de remmende respons. Men merkte ook op dat na verzwakking er een toename was van de excitatoire respons. De controle voor dit experiment bestond uit het testen op verzwakking wanneer een hyperpolarisatiestroom deze spanningsgevoelige kanalen activeerde. Geconcludeerd werd dat verzwakking wordt veroorzaakt door de opening van synaptische receptorkanalen en niet door hyperpolarisatie.
Conclusie
Op elk gegeven moment ontvangt een neuron meerdere inputs van meerdere verschillende neuronen.
Deze inputs kunnen excitatoir of inhibitoir zijn, maar om te bepalen of de drempelpotentiaal zal worden bereikt en een actiepotentiaal zal worden opgewekt of niet, moet naar een aantal factoren worden gekeken.
De verschillende neurotransmitters hebben verschillende effecten, excitatoire neurotransmitters zoals glutamaat veroorzaken het openen van natriumionkanalen waardoor natriumionen instromen en de membraanpotentiaal stijgt. Inhibitoire neurotransmitters zoals GABA kunnen chloride-ionkanalen openen waardoor chloride-ionen instromen of ze kunnen kalium-ionkanalen openen waardoor kalium-ionen uitstromen.
Beide effecten verlagen het membraanpotentiaal, waardoor het van de drempelspanning afwijkt en de kans op het ontstaan van een actiepotentiaal afneemt.
Een andere factor die de kans op het ontstaan van een actiepotentiaal vergroot, is hoe dicht de dendritische ingang zich bij de axonheuvel bevindt; hoe dichter bij de axonheuvel, hoe groter de kans op het ontstaan van een actiepotentiaal. Een axonheuvel is een deel van het soma van een neuron dat het verbindt met het axon.
Excitatoire neurotransmitters verhogen de kans op depolarisatie, de verandering in de membraanspanning wordt een excitatoire postsynaptische potentiaal (EPSP) genoemd. Inhibitoire neurotransmitters verhogen de kans op hyperpolarisatie, deze verandering in de membraanpotentiaal wordt een inhibitoire postsynaptische potentiaal (IPSP) genoemd.
Of de drempelspanning zal worden bereikt, en een actiepotentiaal zal worden geïnitieerd of niet, hangt af van de algebraïsche sommatie van de afzonderlijke ingangen. Er zijn twee soorten optelling.
Sommering van meerdere ingangen van meerdere verschillende presynaptische neuronen die een actiepotentiaal opwekken, wordt ruimtelijke sommatie genoemd. Een hoge frequentie van ingangen van één presynaptisch neuron die worden samengevoegd om een actiepotentiaal te produceren, wordt temporele sommatie genoemd. De duur van deze postsynaptische potentiaal is langer dan de duur tussen de respectieve actiepotentialen.