1. Morphologie
2. Basalschicht
3. Spinöse Schicht
4. Granulöse Schicht
5. Corneum stratum
6. Funktionen
Keratinozyten sind die Hauptzellbestandteile der Epidermis. Sie geben nur wenig extrazelluläre Matrix ab, so dass die Plasmamembranen benachbarter Keratinozyten sehr dicht beieinander liegen. Der Zusammenhalt der Keratinozyten wird durch viele Desmosomen verstärkt, die die Integrität der Epidermis aufrechterhalten. Keratine, eine Art Intermediärfilament des Zytoskeletts, bilden die wichtigste Familie von Proteinen, die von Keratinozyten synthetisiert werden. Keratinozyten durchlaufen einen charakteristischen Lebenszyklus, der in der Basalschicht der Epidermis beginnt und in der oberflächlicheren oder freien Oberfläche der Epidermis endet.
Morphologie und Differenzierung
Die Morphologie der Keratinozyten verändert sich während des Lebenszyklus, der beim Menschen etwa 1 Monat dauert. Die Veränderungen sind progressiv von der Basalschicht, wo die Keratinozyten geboren werden, bis zur oberflächlichen Schicht, wo sie absterben und sich von der Epidermis ablösen. Die morphologischen Unterschiede der Keratinozyten ergeben die Merkmale der Epidermisschichten. Die Schichten oder Strata (Stratum im Singular) sind das Stratum basale, Stratum spinosum, Stratum granulosum und Stratum corneum. Die Schichten können sich je nach Körperregion in ihrer Dicke verändern (Abbildung 1). So kann die Dicke der Epidermis, also die Dicke der Schichten, von 50 µm in Körperregionen mit geringer mechanischer Beanspruchung bis zu 1 mm in solchen Regionen wie Handflächen und Fußsohlen reichen, in denen die Reibung höher ist.

Stratum basale

Das Stratum basale oder Basalschicht (Abbildungen 2 und 4) ist eine eindicke Zellschicht, die hauptsächlich aus Keratinozyten besteht, die sich im tieferen Teil der Epidermis befinden. Diese Zellen sind durch Hemidesmosomen mit der Basallamina verbunden. Diese Verbindung hilft, die Integrität des Epithels zu erhalten und steuert die Proliferation und Differenzierung der Keratinozyten.
Keratinozyten-Stammzellen sind im Stratum basale zu finden. Sie sind rundlich bis säulenförmig, etwa 6-10 µm groß und zeigen aufgrund ihres hohen Gehalts an Ribosomen eine höhere Affinität zu basischen Farbstoffen als differenzierte Keratinozyten. Die Stammzellen der interfollikulären Epidermis differenzieren sich in Keratinozyten, nicht aber in andere epidermale Zelltypen. Auch in den Haarfollikeln gibt es Keratinozyten-Stammzellen, die in Zellen des Haarfollikels (Keratinozyten und Talgdrüsen) und in Keratinozyten der interfollikulären Epidermis differenzieren.
Die Proliferations- und Differenzierungsraten der Keratinozyten-Stammzellen und die Replamationsrate der reifen Keratinozyten müssen im Epithel ähnlich sein. Die Replikationsrate hängt von den Umständen wie Wundheilung, Reibung oder Organwachstum ab. Der Kontakt der Stammzellen mit der Basallamina scheint für die Regulierung dieser Proliferationsrate wichtig zu sein. Wenn zum Beispiel Integrine, Adhäsionsproteine, die Keratinozyten an die Basallamina binden, den Kontakt zur Basallamina verlieren, beginnen die Stammzellen zu differenzieren und in die oberen Schichten der Epidermis zu wandern.
Keratinozyten des Stratum basale synthetisieren einen Teil der Basallamina. Integrine, die Keratinozyten an die Basallamina binden, sind auch an der Aufrechterhaltung der Integrität der Basallamina beteiligt.
Stratum spinosum
Das Stratum spinosum befindet sich direkt über dem Stratum basale (Abbildungen 2 und 4). Es besteht aus mehr oder weniger polyedrischen Keratinozyten von etwa 10 bis 15 µm Größe, größer als die im Stratum basale, mit mehr eosinophilem Zytoplasma und einem oder zwei deutlich sichtbaren Kernen. Im Zytoplasma sind viele Keratinbündel vorhanden, die als Tonofilamente bezeichnet werden und sich zu Tonofibrillen zusammenschließen. Tonofibrillen können lichtmikroskopisch beobachtet werden und tragen zur zytoplasmatischen Färbung mit Eosin bei. In diesem Stratum werden die Keratinozyten als stachelige Keratinozyten bezeichnet, weil sie viele Desmosomen (Abbildung 3) und adhärente Junctions aufweisen. Diese Zell-Zell-Verbindungen sind radial in der Plasmamembran verteilt und werden durch Tonofibrillen verbunden. In der Lichtmikroskopie sehen die Zellverbindungen wie Stacheln aus, daher der Name dieser Schicht.
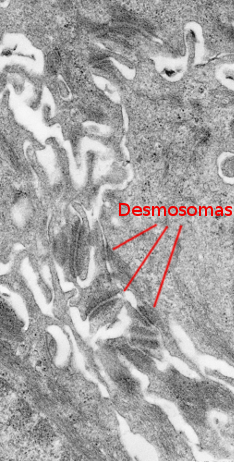
Keratinozyten exprimieren die Intermediärfilament-Keratine im Zytoplasma. Der Typ des exprimierten Keratins ändert sich, wenn sich die Keratinozyten in Richtung der oberen Oberfläche des Epithels bewegen. Im oberflächlicheren Teil des Stratum spinosum werden die Keratinozyten flacher in ihrer Form, was wahrscheinlich durch die fortschreitende Veränderung des Typs der synthetisierten Keratine verursacht wird.
Das Stratum spinosum ist in den Hautregionen dicker, die Reibung oder mechanischen Dehnungen ausgesetzt sind. Die starke Zell-Zell-Adhäsion in diesem Stratum trägt dazu bei, dass die Epidermis widerstandsfähiger wird.
Stratum granulosum
In den äußeren Schichten des Stratum granulosum verändern die Keratinozyten die Genexpression und produzieren Kerato-Hyalin-Granula (Abbildungen 2 und 4). Das Stratum granulosum ist relativ dünn, etwa 15 µm, und die Keratinozyten sind abgeflacht und zeigen viele Kerato-Hyalin-Granula im Zytoplasma. Diese Granula sind polygonal, nicht durch eine Membran abgegrenzt und stark basophil. Sie wachsen progressiv, verstärken die epidermale Barriere und sind der Ausgangspunkt für die Verhornung oder Keratinisierung der oberen Epidermisschichten.

Im Stratum granulosum werden Lamellenkörper beobachtet. Diese Zellstrukturen sind im Stratum basale nicht vorhanden und beginnen im Stratum spinosum sichtbar zu werden. Im Stratum granulosum sind sie sehr zahlreich vorhanden. Lamellenkörperchen sind etwa 100 bis 300 nm große Organellen, die aus Stapeln von Lipidschichten bestehen, also eine lamellare Struktur aufweisen. Ihre Funktion wird im Stratum corneum ausgeübt und steht im Zusammenhang mit dem abgelösten Zellprozess in diesem Stratum und mit der Dichtigkeit der Epidermis. Lamellarkörper werden exozytiert und ihre Bestandteile werden extrazellulär modifiziert, so dass sie parallel zur Hautoberfläche Lipidschichten bilden.
Der Zellkern und die Organellen der Zellen im Stratum granulosum sind degradiert, während das Zytoplasma fast vollständig mit Keratinfilamenten gefüllt ist. In den letzten Stadien der Zelldifferenzierung ist die Durchlässigkeit für Calcium und andere Ionen erhöht. Die Ionen fördern die Aktivierung von Transglutaminasen und führen zur Bildung einer Proteinstruktur unterhalb der Plasmamembran, die als Zellhülle bezeichnet wird. Diese Struktur nimmt mit der Entartung der Keratinozyten an Dicke zu und ist wichtig für die Organisation der Keratinfilamente parallel zur Epitheloberfläche und für die Verbindung dieser Filamente mit den extrazellulären Lamellenkörpern.
Die zwischen den Keratinozyten des Stratum granulosum gebildeten Tight Junctions bilden eine dichte Barriere, die nur von kleinen Molekülen und Ionen durchquert werden kann. Keratinozyten sterben durch einen Prozess ab, der als Verhornung bekannt ist, und werden zu einer verhornten anukleären Zelle. Die Verhornung ist ein anderer Mechanismus als die Apoptose. Bevor sie absterben, synthetisieren Keratinozyten Proteine und Lipide, die zur Bildung der oberflächlichen Barriere der Epidermis beitragen.
Das Stratum lucidum ist eine Schicht aus Keratinozyten zwischen dem Stratum granulosum und dem Stratum corneum. Es befindet sich in Hautregionen mit starker mechanischer Beanspruchung, wie den Fußsohlen.
Stratum corneum
Das Stratum corneum besteht aus entarteten Keratinozyten, den sogenannten Korneozyten (Abbildung 5). Die Korneozyten sind durch Desmosomen (Korneosomen) miteinander verbunden und in eine extrazelluläre Matrix eingebettet, die mit unpolaren Lipiden angereichert und in Schichten organisiert ist. In den letzten Stadien verlieren die Korneozyten ihren Kern, werden mit Keratinen gefüllt und sind von einer Schicht aus Proteinen sowie einer Hülle aus chemisch gebundenen Lipiden umgeben.

Die Dicke des Stratum corneum ist variabel und hängt von der allgemeinen Dicke der Epidermis ab. Je mehr Keratinozyten im Stratum basale gebildet werden, desto dicker ist das Stratum spinosum und das Stratum corneum. Das Absterben von Zellen im Stratum granulosum führt zum Verlust von Desmosomen entlang des Stratum corneum, wodurch die Zell-Zell-Verbindungen schwächer werden und sich daher Zellen und Zellfragmente von der Epidermis ablösen. Filaggrin und Keratin machen 80 bis 90 % der Proteine des Stratum corneum aus.
Diese Schicht schützt vor Austrocknung, Krankheitserregern sowie chemischen und physikalischen Schäden. Wichtig für die Epidermis ist die Verarbeitung der Lipide des Lamellenkörpers. Zum Beispiel wird Glycerin aus Lamellenkörpern gebildet und hilft der Epidermis, die Hydratation aufrechtzuerhalten. Freie Lipide der Oberfläche tragen dazu bei, die Epidermis ein wenig sauer zu halten, mit einem pH-Wert um 5 beim Menschen. Außerdem wird Cholesterinsulfat in Cholesterin umgewandelt, das an der Ablösung der oberflächlicheren Schichten des Stratum corneum beteiligt ist. Die freigesetzten Lipide sind in mehreren Schichten organisiert, die die Undurchlässigkeit der Epidermis ermöglichen.
Funktionen
Keratinozyten bilden die Epidermis der Haut und ihre Hauptfunktion ist es, eine Barriere zwischen der äußeren und der inneren Umgebung zu organisieren. Sie schützen vor mechanischen Schäden, ultraviolettem Licht, Krankheitserregern und schädlichen chemischen Substanzen. Sehr wichtig ist die Rolle der Epidermis, den Wasserverlust zu verhindern. Außerdem sind die Keratinozyten bei der Reparatur nach einer Wunde sehr aktiv.
Wasserverlust-Barriere
Die Verhinderung von Wasserverlust beruht hauptsächlich auf dem Stratum corneum. Es ist wie eine Mauer mit Brüchen und Mörtel, wobei die Brüche die Zellen und der Mörtel die extrazellulären Lipidaggregate sind.
Barriere gegen Krankheitserreger
Die Epidermis ist die erste physikalische Hürde, die Krankheitserreger überwinden müssen, um in den Organismus zu gelangen. Darüber hinaus setzen Keratinozyten Zytokine frei, die Krankheitserreger hemmen und neutralisieren. Nach einer Wunde oder Verletzung stimulieren Keratinozyten die Entzündung und die Aktivierung von Lagerhans-Zellen, den antigenpräsentierenden Zellen der Epidermis.
Hautpigmentierung
Die Pigmentierung der Haut dient dem Schutz vor ultraviolettem Licht. Bei Säugetieren ist Melanin das Pigment, das die Haut dunkler macht. Melanin wird von Melanozyten synthetisiert, einem sehr seltenen Zelltyp, der verstreut im Stratum basale der Epidermis und in den Haarfollikeln zu finden ist. Melanin wird in Organellen im Zytoplasma verpackt, die Melanosomen genannt werden. Die Produktion und Freisetzung von Melanosomen wird sowohl durch Umweltvariablen als auch durch parakrine Substanzen, die von Keratinozyten freigesetzt werden, beeinflusst. Bevor sie freigesetzt werden, werden die Melanosomen durch das Zytoplasma der Melonozyten und durch Zellfortsätze, die sich zwischen den umgebenden Keratinozyten erstrecken, verteilt. Die endgültige Farbe der Haut hängt von der Produktion der Melanosomen und ihrer Verteilung durch die Epidermis ab. Nach einer hohen Melaninsynthese werden Melanosomen aus Melanozyten exozytiert und von benachbarten Keratinozyten aufgenommen. Es wird geschätzt, dass jeder Melanozyt etwa 45 Keratinozyten mit Melanin versorgt. Sobald Melanosomen in die Keratinozyten des Stratum basale aufgenommen wurden, werden sie in die oberen Schichten der Epidermis verlagert, wenn beladene Keratinozyten das Stratum basale verlassen und sich zu den oberen Schichten differenzieren. Auf diese Weise haben alle Epidermisschichten Melanin, auch das Stratum corneum. Insgesamt hängt die Farbe der Haut von der Menge an Melanin ab, die Melanozyten und Keratinozyten enthalten.
Wundheilung
Nach einer Wunde wechseln die Keratinozyten von einem stationären Zustand in einen anderen, in dem sie sich zu bewegen beginnen. Die Ablösung von der Basalmembran, vermittelt durch den Verlust von Desmosomen und die Veränderung der Expression von Integrinen, lässt die Keratinozyten beweglich werden, ebenso wie die Reaktion auf Zytokine, die von anderen Zellen freigesetzt werden. Auf diese Weise können Keratinozyten zu den geschädigten Bereichen wandern und diese abdichten.
Bibliographie
Eckert RL, Rorke EA. 1989. Molekularbiologie der Keratinozytendifferenzierung. Environmental health perspectives 80:109-116.
Ginnell F. 1992. Wundreparatur, Keratinozytenaktivierung und Integrinmodulation. Journal of cell sciences 101:1-5.
Madison KC. 2003. Barrierefunktion der Haut: „La Raison d’EŒtre'“ der Epidermis. The journal of investigative dermatology 121: 231-241.
Nemes Z, Steinert PM. 1999. Ziegel und Mörtel der epidermalen Barriere. Experimental and molecular medicine 1: 5-19.
Proksch E, Brandner JM, Jensen J-M. 2008. Die Haut: eine unverzichtbare Barriere. Experimental Dermatology 17: 1063-1072.
Simpson CL, Patel DM, Green KJ. 2011. Deconstructing the skin: cytoarchitectural determinants of epidermal morphogenesis. Nature Reviews in Molecular and Cell Biology 12:568-580.
Yamaguchi Y, Brenner M, Hearing VJ. 2007. The Regulation of Skin Pigmentation. Journal of biological chemistry 282:27557-27561.