1. Morphologie
2. strate basale
3. strate épineuse
4. strate granuleuse
5. Stratum cornéen
6. Fonctions
Les kératinocytes sont les principaux composants cellulaires de l’épiderme. Ils libèrent très peu de matrice extracellulaire de sorte que la membrane plasmique des kératinocytes adjacents est très proche. La cohésion des kératinocytes est renforcée par de nombreux desmosomes qui maintiennent l’intégrité de l’épiderme. Les kératines, un type de filament intermédiaire du cytosquelette, constituent la principale famille de protéines synthétisées par les kératinocytes. Les kératinocytes suivent un cycle de vie caractéristique qui commence dans la couche basale de l’épiderme et se termine dans la surface plus superficielle ou libre de l’épiderme.
Morphologie et différenciation
La morphologie des kératinocytes change au cours du cycle de vie, qui dure environ 1 mois chez l’homme. Les changements sont progressifs depuis la couche basale, où les kératinocytes sont nés, jusqu’à la couche superficielle, où ils meurent et se détachent de l’épiderme. Les différences morphologiques des kératinocytes produisent les caractéristiques des couches épidermiques. Les couches ou strates (stratum au singulier) sont le stratum basale, le stratum spinosum, le stratum granulosum et le stratum corneum. L’épaisseur de ces couches peut varier en fonction de la région du corps (figure 1). Ainsi, l’épaisseur de l’épiderme, c’est-à-dire l’épaisseur des strates, peut aller de 50 µm dans les régions du corps où les contraintes mécaniques sont faibles à 1 mm dans les régions comme la paume des mains et la plante des pieds, où les frottements sont plus importants.

Stratum basale

La couche basale ou couche basale (Figures 2 et 4) est une couche cellulaire épaisse composée principalement de kératinocytes que l’on trouve dans la partie profonde de l’épiderme. Ces cellules sont attachées à la lame basale par des hémidesmosomes. Cette connexion permet de conserver l’intégrité de l’épithélium et de contrôler la prolifération et la différenciation des kératinocytes.
Les cellules souches kératinocytaires se trouvent dans le stratum basale. Elles sont de forme arrondie à colonnaire, d’une taille d’environ 6 à 10 µm, et présentent une plus grande affinité pour les colorants basiques que les kératinocytes différenciés en raison de leur teneur élevée en ribosomes. Les cellules souches de l’épiderme interfolliculaire se différencient en kératinocytes, mais pas en d’autres types de cellules épidermiques. Il existe également des cellules souches kératinocytaires dans les follicules pileux qui se différencient en cellules du follicule pileux (kératinocytes et glandes sébacées) et en kératinocytes de l’épiderme interfolliculaire.
Les taux de prolifération et de différenciation des cellules souches kératinocytaires et le taux de remplacement des kératinocytes matures doivent être similaires dans l’épithélium. Le taux de remplacement dépend de circonstances telles que la réparation de la plaie, le frottement ou la croissance de l’organe. Le contact des cellules souches avec la lame basale semble être important pour réguler ce taux de prolifération. Par exemple, lorsque les intégrines, protéines d’adhésion qui fixent les kératinocytes à la lame basale, perdent le contact avec cette dernière, les cellules souches commencent à se différencier et à migrer vers les couches supérieures de l’épiderme.
Les kératinocytes du stratum basale synthétisent une partie de la lame basale. Les intégrines qui fixent les kératinocytes à la lame basale sont également impliquées dans le maintien de l’intégrité de la lame basale.
Stratum spinosum
Le stratum spinosum se trouve juste au-dessus du stratum basale (figures 2 et 4). Il est constitué de kératinocytes plus ou moins polyédriques d’environ 10 à 15 µm, plus grands que ceux du stratum basale, avec un cytoplasme plus éosinophile et un ou deux noyaux bien visibles. De nombreux faisceaux de kératine sont présents dans le cytoplasme, appelés tonofilaments, qui s’assemblent pour former des tonofibrilles. Les tonofibrilles peuvent être observées en microscopie optique et contribuent à la coloration cytoplasmique à l’éosine. Dans cette strate, les kératinocytes sont appelés kératinocytes épineux car ils partagent de nombreux desmosomes (Figure 3) et des jonctions adhérentes. Ces jonctions cellule-cellule sont distribuées radialement dans la membrane plasmique et sont reliées par des tonofibrilles. En microscopie optique, les jonctions cellulaires ressemblent à des épines, d’où le nom de cette couche.
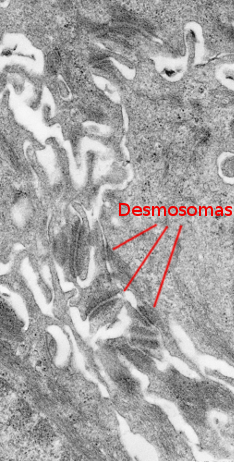
Les kératinocytes expriment les kératines des filaments intermédiaires dans le cytoplasme. Le type de kératine exprimé change à mesure que les kératinocytes se déplacent vers la surface supérieure de l’épithélium. Dans la partie plus superficielle du stratum spinosum, les kératinocytes prennent une forme plus aplatie, probablement induite par le changement progressif du type de kératines synthétisées.
La stratum spinosum est plus épaisse dans les régions de la peau exposées aux frottements ou aux étirements mécaniques. La forte adhésion cellule-cellule dans cette strate permet d’obtenir un épiderme plus résistant.
Stratum granulosum
Dans les couches externes du stratum granulosum, les kératinocytes modifient l’expression génétique et produisent des granules de kérato-hyaline (Figures 2 et 4). Le stratum granulosum est relativement mince, environ 15 µm, et les kératinocytes sont aplatis et montrent de nombreux granules de kérato-hyaline dans le cytoplasme. Ces granules sont polygonaux, non délimités par une membrane, et fortement basophiles. Ils croissent progressivement, rendent plus forte la barrière épidermique et sont le point de départ de la cornification ou kératinisation des couches supérieures de l’épiderme.

Des corps lamellaires sont observés dans le stratum granulosum. Ces structures cellulaires ne sont pas présentes dans le stratum basale, et commencent à être visibles dans le stratum spinosum. Elles sont très abondantes dans le stratum granulosum. Les corps lamellaires sont des organites d’une taille d’environ 100 à 300 nm, constitués d’empilements de couches lipidiques, ils présentent donc une structure lamellaire. Leur fonction s’exerce dans la couche cornée et est liée au processus de détachement cellulaire dans cette couche et à l’imperméabilité de l’épiderme. Les corps lamellaires sont exocytés et leurs composants sont modifiés de manière extracellulaire pour former des couches lipidiques parallèles à la surface de la peau.
Le noyau et les organites des cellules du stratum granulosum sont dégradés, tandis que le cytoplasme est presque plein de filaments de kératine. La perméabilité au calcium et à d’autres ions est accrue au cours des dernières étapes de la différenciation cellulaire. Les ions stimulent l’activation des transglutaminases et conduisent à la formation d’une structure protéique sous la membrane plasmique, appelée enveloppe cellulaire. Cette structure croît en épaisseur au fur et à mesure que les kératinocytes dégénèrent et elle est importante pour organiser les filaments de kératine parallèlement à la surface épithéliale et pour combiner ces filaments avec les corps lamellaires extracellulaires.
Les jonctions serrées formées entre les kératinocytes du stratum granulosum constituent une barrière étanche qui ne peut être traversée que par de petites molécules et des ions. Les kératinocytes meurent par un processus appelé cornification, et deviennent une cellule anucléaire cornifiée. La cornification est un mécanisme différent de l’apoptose. Avant de mourir, les kératinocytes synthétisent des protéines et des lipides qui contribuent à la formation de la barrière superficielle de l’épiderme.
Le stratum lucidum est une couche de kératinocytes située entre le stratum granulosum et le stratum corneum. On le retrouve les régions de la peau soumises à de fortes contraintes mécaniques, comme la plante des pieds.
Stratum corneum
Le stratum corneum est constitué de kératinocytes dégénérés, les fameux cornéocytes (figure 5). Les cornéocytes sont reliés les uns aux autres par des desmosomes (cornéosomes) et noyés dans une matrice extracellulaire enrichie en lipides non polaires organisée en couches. Au cours des derniers stades, les cornéocytes perdent leur noyau, se remplissent de kératines et sont entourés d’une couche de protéines, ainsi que d’une couverture de lipides liés chimiquement.

L’épaisseur du stratum corneum est variable et dépend de l’épaisseur générale de l’épiderme. Ainsi, plus les kératinocytes sont produits dans la couche basale, plus l’épaisseur est importante dans les couches spinosum et cornée. La mort des cellules dans le stratum granulosum entraîne la perte des desmosomes le long du stratum corneum, ce qui rend les jonctions cellule-cellule plus faibles et donc les cellules et les fragments de cellules se détachent de l’épiderme. La filaggrine et la kératine représentent 80 à 90 % des protéines du stratum corneum.
Cette couche protège du dessèchement, des agents pathogènes et des dommages chimiques et physiques. La transformation des lipides des corps lamellaires est importante pour l’épiderme. Par exemple, le glycérol est formé à partir des corps lamellaires et aide l’épiderme à maintenir son hydratation. Les lipides libres de la surface contribuent à maintenir l’épiderme un peu acide, avec un pH d’environ 5 chez l’homme. De plus, le sulfate de cholestérol est transformé en cholestérol, ce qui participe au décollement des couches les plus superficielles de la couche cornée. Les lipides libérés sont organisés en plusieurs feuillets qui rendent possible l’imperméabilité de l’épiderme.
Fonctions
Les kératinocytes forment l’épiderme de la peau et leur principale fonction est d’organiser une barrière entre le milieu extérieur et le milieu intérieur. Ils protègent contre les dommages mécaniques, les rayons ultraviolets, les agents pathogènes et les substances chimiques nocives. Le rôle de l’épiderme est très important pour prévenir la perte d’eau. De plus, les kératinocytes sont très actifs lors de la réparation après une blessure.
Barrière de la perte d’eau
La prévention de la perte d’eau repose principalement sur la couche cornée. C’est comme un mur avec des cassures et du mortier, où les cassures sont les cellules et le mortier les agrégats lipidiques extracellulaires.
Barrière contre les agents pathogènes
L’épiderme est le premier obstacle physique que les agents pathogènes doivent franchir pour pénétrer dans l’organisme. En outre, les kératinocytes libèrent des cytokines qui inhibent et neutralisent les agents pathogènes. Après une blessure ou un dommage, les kératinoncytes stimulent l’inflammation et l’activation des cellules de Lagerhans, qui sont les cellules présentatrices d’antigènes de l’épiderme.
Pigmentation de la peau
La pigmentation de la peau a pour but de protéger contre les rayons ultraviolets. Chez les mammifères, la mélanine est le pigment qui rend la peau plus foncée. La mélanine est synthétisée par les mélanocytes, un type cellulaire très rare que l’on trouve dispersé dans la couche basale de l’épiderme et dans les follicules pileux. La mélanine est emballée dans des organelles du cytoplasme appelées mélanosomes. La production et la libération des mélanosomes sont influencées par des variables environnementales ainsi que par des substances paracrines libérées par les kératinocytes. Avant leur libération, les mélanosomes sont distribués par le cytoplasme des melonocytes et les processus cellulaires qui s’étendent parmi les kératinocytes environnants. La couleur finale de la peau dépend de la production de mélanosomes et de leur distribution dans l’épiderme. Après une forte synthèse de mélanine, les mélanosomes sont exocytés par les mélanocytes et capturés par les kératinocytes voisins. On estime que chaque mélanocyte fournit de la mélanine à environ 45 kératinocytes. Une fois que les mélanosomes ont été incorporés dans les kératinocytes de la couche basale, ils sont déplacés vers les strates supérieures de l’épiderme lorsque les kératinocytes chargés quittent la couche basale et se différencient pour former les strates supérieures. De cette façon, toutes les strates de l’épiderme ont de la mélanine, même la couche cornée. Au total, la couleur de la peau dépend de la quantité de mélanine que contiennent les mélanocytes et les kératinocytes.
Réparation des blessures
Après une blessure, les kératinocytes passent d’un état stable à un autre où ils commencent à bouger. La déconnexion de la membrane basale médiée par la perte des desmosomes et le changement dans l’expression des intégrines laissent à la motilité des kératinocytes, ainsi que la réponse aux cytokines libérées par d’autres cellules. De cette façon, les kératinocytes peuvent se déplacer vers les zones endommagées et les sceller.
Bibliographie
Eckert RL, Rorke EA. 1989. Biologie moléculaire de la différenciation des kératinocytes. Perspectives de la santé environnementale 80:109-116.
Grinnell F. 1992. Réparation des plaies, activation des kératinocytes et modulation des intégrines. Journal of cell sciences 101:1-5.
Madison KC. 2003. La fonction de barrière de la peau : « La Raison d’EŒtre » de l’Epiderme. The journal of investigative dermatology 121 : 231-241.
Nemes Z, Steinert PM. 1999. Les briques et le mortier de la barrière épidermique. Médecine expérimentale et moléculaire 1 : 5-19.
Proksch E, Brandner JM, Jensen J-M. 2008. La peau : une barrière indispensable. Dermatologie expérimentale 17 : 1063-1072.
Simpson CL, Patel DM, Green KJ. 2011. Deconstructing the skin : cytoarchitectural determinants of epidermal morphogenesis. Nature reviews in molecular and cell biology 12:568-580.
Yamaguchi Y, Brenner M, Hearing VJ. 2007. La régulation de la pigmentation de la peau. Journal of biological chemistry 282:27557-27561.