Pores nucléaires
L’enveloppe nucléaire est perforée de minuscules trous appelés pores nucléaires, dont la découverte remonte au milieu du XXe siècle. Ces pores régulent le passage des molécules entre le noyau et le cytoplasme, permettant à certaines de traverser la membrane, mais pas à d’autres. Les éléments constitutifs de la fabrication de l’ADN et de l’ARN font partie des matériaux qui sont autorisés à pénétrer dans le noyau, tout comme les molécules qui fournissent l’énergie nécessaire à la construction du matériel génétique. Les sous-unités ribosomales, qui sont construites dans les nucléosomes, sont un excellent exemple de matériaux qui doivent être autorisés à quitter le noyau et à entrer dans le cytoplasme.
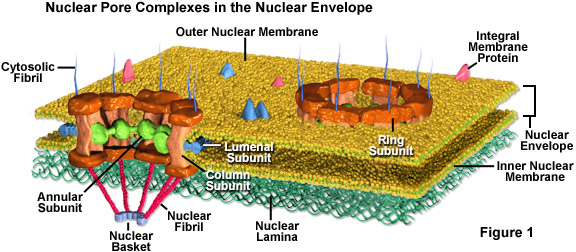
Les pores nucléaires sont entièrement perméables aux petites molécules jusqu’à la taille des plus petites protéines, mais forment une barrière empêchant la plupart des grosses molécules d’entrer dans le noyau. Pourtant, certaines protéines plus grosses, comme les histones, sont admises dans le noyau alors que les pores devraient être trop petits pour les laisser passer. On pense généralement que la structure protéique élaborée appelée complexe de pores nucléaires (voir figure 1) qui entoure chaque pore joue un rôle clé en permettant le transport actif d’un ensemble sélectionné de grosses molécules vers et hors du noyau.
Le complexe de pores nucléaires est composé de plusieurs sous-unités. Autour de l’intérieur du pore se trouve un matériau non membraneux organisé en un anneau qui prolonge des structures en forme de rayons vers le centre du petit canal. La paroi du pore proprement dite est principalement constituée de sous-unités colonnaires, et les sous-unités luménales, avec l’aide de protéines transmembranaires, ancrent l’ensemble du complexe du pore dans l’enveloppe nucléaire. De plus, de minuscules fibrilles s’étendent généralement des deux côtés du complexe et se rassemblent en configurations en forme de panier sur le côté nucléaire du complexe. La composition protéique de ces fibrilles est différente sur les côtés opposés de la structure.
En plus de leur rôle dans le transport nucléaire, les pores nucléaires sont importants en tant que sites où la membrane externe et la membrane interne de l’enveloppe nucléaire sont fusionnées. En raison de cette fusion, les membranes peuvent être considérées comme continues l’une avec l’autre, bien qu’elles aient des caractéristiques biochimiques différentes et puissent fonctionner de manière distincte. Comme la membrane nucléaire externe est également continue avec la membrane du réticulum endoplasmique (RE), elle et la membrane nucléaire interne peuvent échanger des matériaux membranaires avec le RE. Cette capacité permet à l’enveloppe nucléaire de s’agrandir ou de se réduire lorsque cela est nécessaire pour accueillir le contenu dynamique du noyau.
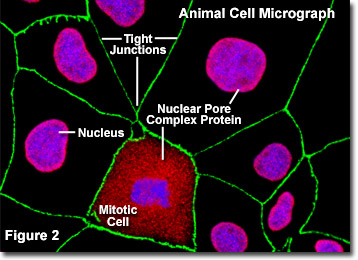
La figure 2 illustre une image numérique par fluorescence d’une culture adhérente de cellules rénales canines de Madin-.Darby (lignée MDCK) colorée avec des sondes fluorescentes ciblant le noyau (bleu), les protéines du complexe des pores nucléaires (rouge) et les jonctions serrées formées entre les cellules épithéliales (vert) pour démontrer la proximité de ces structures. Les pores nucléaires de ces cellules ont été ciblés avec un anticorps polyclonal à large spectre dirigé contre une grande famille de protéines du complexe du pore nucléaire, qui constitue un outil utile pour étudier la morphologie et la composition du noyau et de l’enveloppe nucléaire. Le mélange d’anticorps est également utile pour étudier les changements de la structure nucléaire au cours de la mitose et de la méiose (notez la cellule mitotique dans la partie centrale inférieure de la figure 2).
La densité des pores varie fortement et a tendance à être plus importante parmi les cellules fortement activées et différenciées, comme les cellules du foie. Une cellule typique de mammifère présente environ 3 000 à 4 000 pores le long de son enveloppe nucléaire. Les ovocytes de certains amphibiens, cependant, ont des noyaux si grands et une telle densité de pores que l’enveloppe nucléaire d’une des cellules peut contenir plus de dix millions de pores. Par conséquent, ces ovocytes ont été fortement utilisés dans les études des complexes de pores nucléaires et du transport nucléaire.
Retour au noyau cellulaire
Retour à la structure cellulaire animale
Retour à la structure cellulaire végétale
Questions ou commentaires ? Envoyez-nous un courriel.
© 1995-2021 par Michael W. Davidson et l’Université d’État de Floride. Tous droits réservés. Aucune image, graphique, logiciel, script ou applet ne peut être reproduit ou utilisé de quelque manière que ce soit sans l’autorisation des détenteurs des droits d’auteur. L’utilisation de ce site Web signifie que vous acceptez toutes les conditions légales énoncées par les propriétaires.
Ce site Web est maintenu par notre
Graphics & Web Programming Team
en collaboration avec Optical Microscopy at the
National High Magnetic Field Laboratory.
Dernière modification : Vendredi 13 novembre 2015 à 02:18
Compte d’accès depuis le 14 mai 2005 : 157213
Microscopes fournis par :
![]()
![]()