Pori nucleari
L’involucro nucleare è perforato da piccoli fori noti come pori nucleari, che sono stati scoperti per la prima volta a metà del ventesimo secolo. Questi pori regolano il passaggio delle molecole tra il nucleo e il citoplasma, permettendo ad alcune di passare attraverso la membrana, ma non ad altre. I mattoni per la produzione di DNA e RNA sono alcuni dei materiali che possono entrare nel nucleo, così come le molecole che forniscono l’energia per la costruzione del materiale genetico. Le subunità ribosomiali, che sono costruite nei nucleosomi, sono un primo esempio di materiali che devono poter lasciare il nucleo ed entrare nel citoplasma.
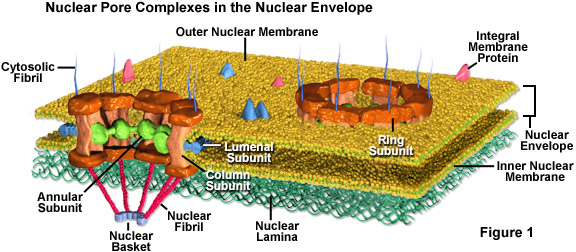
I pori nucleari sono completamente permeabili alle piccole molecole fino alle dimensioni delle proteine più piccole, ma formano una barriera che tiene la maggior parte delle grandi molecole fuori dal nucleo. Eppure, alcune proteine più grandi, come gli istoni, sono ammessi nel nucleo nonostante il fatto che i pori dovrebbero essere troppo piccoli per lasciarli passare. Si pensa generalmente che l’elaborata struttura proteica chiamata complesso dei pori nucleari (vedi Figura 1) che circonda ogni poro svolga un ruolo chiave nel permettere il trasporto attivo di una serie selezionata di grandi molecole dentro e fuori il nucleo.
Il complesso dei pori nucleari è composto da diverse subunità. Circonda l’interno del poro un materiale non membranoso organizzato in un anello che estende strutture a raggiera verso il centro del piccolo canale. La parete effettiva del poro è prevalentemente composta da subunità colonnari, e le subunità lumenali, con l’aiuto di proteine transmembrana, ancorano l’intero complesso del poro nell’involucro nucleare. Inoltre, piccole fibrille di solito si estendono da entrambi i lati del complesso e si riuniscono in configurazioni simili a cesti sul lato nucleare del complesso. La composizione proteica di queste fibrille è diversa sui lati opposti della struttura.
Oltre al loro ruolo nel trasporto nucleare, i pori nucleari sono importanti come siti dove la membrana esterna e la membrana interna dell’involucro nucleare sono fuse insieme. Grazie a questa fusione, le membrane possono essere considerate continue l’una con l’altra anche se hanno caratteristiche biochimiche diverse e possono funzionare in modi diversi. Poiché la membrana nucleare esterna è anche continua con la membrana del reticolo endoplasmatico (ER), sia essa che la membrana nucleare interna possono scambiare materiali membranosi con l’ER. Questa capacità permette all’involucro nucleare di ingrandirsi o rimpicciolirsi quando necessario per accogliere il contenuto dinamico del nucleo.
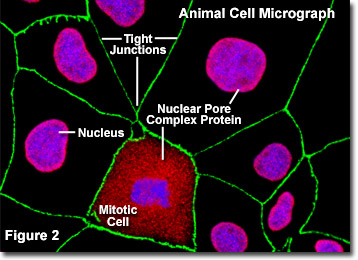
Illustrata nella Figura 2 è un’immagine digitale a fluorescenza di una coltura aderente di cellule di rene canino Madin-Darby canine (linea MDCK) colorate con sonde fluorescenti rivolte al nucleo (blu), le proteine del complesso dei pori nucleari (rosso) e le giunzioni strette formate tra le cellule epiteliali (verde) per dimostrare la vicinanza di queste strutture. I pori nucleari di queste cellule sono stati bersagliati con un anticorpo policlonale ad ampio spettro per una grande famiglia di proteine del complesso del poro nucleare, che serve come strumento utile per studiare la morfologia e la composizione del nucleo e dell’involucro nucleare. La miscela di anticorpi è anche utile per studiare i cambiamenti nella struttura nucleare durante la mitosi e la meiosi (si noti la cellula mitotica nella parte centrale inferiore della Figura 2).
La densità dei pori varia notevolmente e tende ad essere maggiore tra le cellule che sono altamente attive e differenziate, come le cellule del fegato. Una tipica cellula di mammifero presenta circa 3.000 – 4.000 pori lungo il suo involucro nucleare. Gli ovociti di alcuni anfibi, tuttavia, hanno nuclei così grandi e una tale densità di pori che l’involucro nucleare di una delle cellule può contenere più di dieci milioni di pori. Di conseguenza, questi oociti sono stati molto utilizzati negli studi sui complessi dei pori nucleari e sul trasporto nucleare.
RIENTAMENTO AL NUCLEO CELLULARE
RIENTAMENTO ALLA STRUTTURA CELLULARE ANIMALE
RIENTAMENTO ALLA STRUTTURA CELLULARE VEGETALE
Domande o commenti? Inviaci un’e-mail.
© 1995-2021 di Michael W. Davidson e della Florida State University. Tutti i diritti riservati. Nessuna immagine, grafica, software, script o applet può essere riprodotta o utilizzata in qualsiasi modo senza il permesso dei titolari del copyright. L’uso di questo sito web implica l’accettazione di tutti i termini e le condizioni legali stabilite dai proprietari.
Questo sito web è mantenuto dal nostro
Graphics & Web Programming Team
in collaborazione con Optical Microscopy at the
National High Magnetic Field Laboratory.
L’ultima modifica: Friday, Nov 13, 2015 at 02:18 PM
Access Count Since May 14, 2005: 157213
Microscopi forniti da:
![]()
![]()