1. 形態
2.基底層
3.有棘層
4.顆粒層
5.
5. Corneum stratum
6.
ケラチノサイトは、表皮の主要な細胞構成要素です。 このようにしてできたのが「角質細胞」で、細胞外マトリックスをほとんど出さないため、隣り合う角質細胞の細胞膜は非常に接近しています。 ケラチノサイトの結合は、表皮の統合性を維持する多くのデスモソームによって強化されている。 ケラチンは細胞骨格の中間フィラメントの一種であり、ケラチノサイトが合成するタンパク質の主要なファミリーを形成している。 ケラチノサイトは、表皮の基底層で始まり、表皮のより表層または自由表面で終わる特徴的なライフサイクルを経る。
形態と分化
ケラチノサイトの形態は、ヒトの場合、約1ヶ月間のライフサイクルの間に変化します。 その変化は、ケラチノサイトが生まれた基底層から、ケラチノサイトが死滅して表皮から剥離する表層に至るまで、段階的に行われます。 ケラチノサイトの形態的な違いが、表皮の層の特徴を生み出しているのである。 表皮の層とは、基底層、有棘層、顆粒層、角質層のことである。 これらの層は、体の部位によって厚さが変わります(図1)。 したがって、表皮の厚さ、すなわち地層の厚さは、機械的ストレスの少ない身体部位の50μmから、手のひらや足の裏のように摩擦が大きい部位の1mmまでの範囲である可能性があります。

基底層

基底層(図2、4)は、表皮の深部にあるケラチノサイトを中心とした1枚の厚い細胞層です。 これらの細胞は、ヘミデスモソームを介して基底膜に結合している。 この結合は、上皮の完全性を維持するのに役立ち、ケラチノサイトの増殖と分化を制御する。
ケラチノサイトの幹細胞は、基底層に存在します。
ケラチノサイトの幹細胞は、基底層に存在し、丸みを帯びた円柱状で、大きさは約6〜10μm、リボソームを多く含むため、分化したケラチノサイトよりも塩基性色素への親和性が高いことが特徴である。 毛包間表皮の幹細胞はケラチノサイトに分化するが、他の表皮細胞タイプには分化しない。
毛包にもケラチノサイトの幹細胞があり、毛包の細胞(ケラチノサイトと皮脂腺)や毛包間表皮のケラチノサイトに分化します。
ケラチノサイトの幹細胞の増殖率と分化率、そして成熟したケラチノサイトの置換率は、上皮において同様である必要があります。 置換率は、傷の修復、摩擦、臓器の成長などの状況に応じて変化する。 幹細胞の基底膜との接触は、この増殖率の調節に重要であると考えられる。 例えば、ケラチノサイトを基底膜に接着する接着タンパク質であるインテグリンが基底膜との接触を緩めると、幹細胞は分化を始め、表皮の上層に移動するという。
基底層のケラチノサイトは基底膜の一部を合成する。 ケラチノサイトを基底膜に付着させるインテグリンは、基底膜の完全性の維持にも関与している。
有棘層
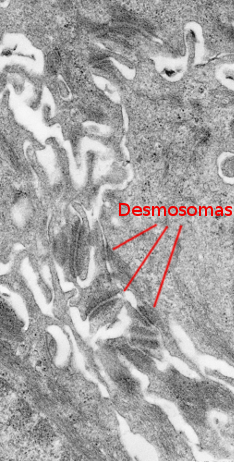
有棘層は、基底層のすぐ上にあります(図2、4)。 多角形のケラチノサイトで構成されており、大きさは10〜15μmで、基底層のケラチノサイトよりも大きく、細胞質は好酸球性で、核は1〜2個はっきりと見える。 細胞質内にはトノフィラメントと呼ばれる多くのケラチン束が存在し、それらが結合してトノフィブリルを形成している。 トノフィブリルは、光学顕微鏡で観察することができ、エオシンによる細胞質の染色に寄与する。 この層では、ケラチノサイトは多くのデスモソーム(図3)と接着接合部を共有しているため、有棘ケラチノサイトと呼ばれる。 これらの細胞間接合部は、細胞膜に放射状に分布し、トノフィブリルによって連結されている。 顕微鏡で見ると、この細胞接合部が棘のように見えるので、この層の名前になっています。
ケラチノサイトは細胞質に中間フィラメントケラチンを発現している。 発現するケラチンの種類は、ケラチノサイトが上皮の上面に向かうにつれて変化する。 これは、合成されるケラチンの種類が徐々に変化しているためと考えられる。
角質層は、皮膚が擦れたり、機械的に伸ばされたりする部分で厚くなっています。 この層では、細胞と細胞の接着が強く、より抵抗力のある表皮を得ることができます。
顆粒層
顆粒層の外側では、ケラチノサイトが遺伝子の発現を変化させ、ケラト・ヒアリン顆粒を生成します(図2、4)。 顆粒層は約15μmと比較的薄く、ケラチノサイトは扁平で、細胞質には多くのケラト・ヒアリン顆粒が見られる。 これらの顆粒は多角形で、膜に囲まれておらず、高度に好塩基性である。 これらの顆粒は徐々に成長し、表皮のバリアーを強化し、表皮上層の角化または角質化の出発点となります。

顆粒層にはラメラ体が観察されます。 これらの細胞構造は、基底層には存在せず、有棘層で見え始めます。 顆粒層では非常に多く見られる。 ラメラ小体は、約100~300nmの大きさのオルガネラで、脂質の層が積み重なってできているため、ラメラ構造をしている。 ラメラ小体の機能は角質層で発揮され、角質層での剥離細胞プロセスや表皮の不透水性に関係している。 ラメラ体は細胞外に放出され、その成分は細胞外で修飾され、皮膚表面に平行な脂質層を形成します。
顆粒層の細胞の核や細胞小器官は分解され、細胞質はケラチンフィラメントでほぼ満たされています。 細胞分化の最終段階では、カルシウムなどのイオンに対する透過性が高まる。 イオンがトランスグルタミナーゼの活性化を促し、細胞膜の下に細胞外被と呼ばれるタンパク質構造が形成される。 この構造は、ケラチノサイトの変性に伴って厚みを増し、ケラチンフィラメントを上皮表面に平行に編成したり、これらのフィラメントを細胞外のラメラ体と結合させるのに重要な役割を果たしています。
顆粒層のケラチノサイトの間に形成されたタイトジャンクションは、低分子やイオンだけが通過できる密閉されたバリアとなっています。 ケラチノサイトは角化と呼ばれる過程で死滅し、角化した無核細胞となる。 角化はアポトーシスとは異なるメカニズムである。 ケラチノサイトは死ぬ前にタンパク質や脂質を合成し、表皮の表層バリアの形成に貢献する。
角質層は、顆粒層と角質層の間にあるケラチノサイトの層です。
角質層
角質層は、変性したケラチノサイト、いわゆる角質細胞で構成されています(図5)。 角質細胞はデスモソーム(コルネオソーム)によって互いに連結され、層状に構成された非極性脂質を豊富に含む細胞外マトリックスに埋め込まれている。 最後の段階では、角質細胞は核を失い、ケラチンでいっぱいになり、タンパク質の層と、化学的に結合した脂質のカバーに囲まれます。

角質層の厚さは可変で、表皮の一般的な厚さに依存します。 基底層で角質細胞が多く作られると、有棘層や角質層の厚みが増します。 顆粒層の細胞が死滅すると、角質層に沿ってデスモソームが消失し、細胞間の結合が弱くなるため、細胞や細胞片が表皮から剥離する。 フィラグリンとケラチンは、角質層のタンパク質の80〜90%を占めています。
この層は、乾燥や病原菌、化学的・物理的なダメージから守る役割を果たしています。 表皮にとって重要なのは、ラメラ体の脂質の処理である。 例えば、グリセロールはラメラ体から形成され、表皮の水分維持を助けます。 また、表面の遊離脂質は、表皮をやや酸性に保つことに貢献しており、ヒトではpH5程度である。 さらに、硫酸コレステロールはコレステロールに変化し、角質層のより表層の剥離に関与している。 放出された脂質はいくつかのシートにまとめられ、表皮の不透水性を可能にします。
機能
ケラチノサイトは皮膚の表皮を形成し、その主な機能は外部環境と内部環境の間のバリアを形成することです。 ケラチノサイトは、機械的損傷、紫外線、病原体、有害な化学物質などから皮膚を保護します。 表皮の役割としては、水分の損失を防ぐことが非常に重要である。 また、ケラチノサイトは、傷を修復する際にも非常に活躍します。
水のバリア
水の流出を防ぐには、主に角質層に頼ることになります。 角質層は、細胞と細胞外の脂質の集合体で構成されており、壁のようなものです。
病原体に対するバリアー
表皮は、病原体が生体内に侵入するための最初の物理的ハードルです。 また、ケラチノサイトは、病原体を抑制・中和するサイトカインを放出します。
皮膚の色素沈着
皮膚の色素沈着は、紫外線から身を守るためのものです。 哺乳類では、皮膚を黒くする色素はメラニンです。 メラニンは、表皮の基底層や毛包に散在するごく少数の細胞であるメラノサイトによって合成される。 メラニンは、メラノソームと呼ばれる細胞質内の小器官にパッケージされる。 メラノソームの生成と放出は、環境変数やケラチノサイトから放出されるパラクリン物質に影響される。 メラノソームは、放出される前に、メラノサイトの細胞質や周囲のケラチノサイトに伸びる細胞突起を介して分配される。 皮膚の最終的な色は、メラノソームの生成と表皮内の分布に依存する。 メラニンが大量に合成されると、メラノサイトからメラノソームが放出され、隣接するケラチノサイトに取り込まれる。 1つのメラノサイトが約45のケラチノサイトにメラニンを供給すると推定されている。 メラノソームが基底層のケラチノサイトに取り込まれると、積み込まれたケラチノサイトが基底層から離れて上層に分化する際に、メラノソームは上層の表皮に移動する。 このようにして、表皮のすべての層、つまり角質層にまでメラニンが存在するようになる。 つまり、皮膚の色は、メラノサイトやケラチノサイトが持つメラニンの量によって決まるのです。
傷の修復
傷ができた後、ケラチノサイトは定常状態から動き出す状態に変化します。 デスモソームの消失による基底膜からの剥離やインテグリンの発現変化により、ケラチノサイトの運動性が向上し、他の細胞から放出されるサイトカインにも反応するようになる。 このようにして、ケラチノサイトはダメージを受けた部分に移動し、封鎖することができる。
参考文献
Eckert RL, Rorke EA. 1989. Molecular biology of keratinocyte differentiation(ケラチノサイト分化の分子生物学)。 Environmental Health Perspectives 80:109-116.
Grinnell F. 1992. 傷の修復、ケラチノサイトの活性化とインテグリンの調節。 Journal of cell sciences 101:1-5.
Madison KC. 2003. 皮膚のバリア機能。 “表皮の「La Raison d’EŒtre」。 The journal of investigative dermatology 121: 231-241.
Nemes Z, Steinert PM. 1999. Bricks and mortar of the epidermal barrier (表皮バリアのレンガとモルタル). Experimental and molecular medicine 1: 5-19.
Proksch E, Brandner JM, Jensen J-M. 2008. 皮膚:必要不可欠なバリア。 Experimental dermatology 17: 1063-1072.
Simpson CL, Patel DM, Green KJ. 2011. Deconstructing the skin: cytoarchitectural determinants of epidermal morphogenesis. Nature reviews in molecular and cell biology 12:568-580.
Yamaguchi Y, Brenner M, Hearing VJ. 2007. The Regulation of Skin Pigmentation.
山口由美子さんは、2007年に「The Regulation of Skin Pigment」を発表しました。