Kernporen
Die Kernhülle ist mit winzigen Löchern perforiert, die als Kernporen bezeichnet werden und erst Mitte des zwanzigsten Jahrhunderts entdeckt wurden. Jahrhunderts entdeckt wurden. Diese Poren regulieren den Durchgang von Molekülen zwischen dem Kern und dem Zytoplasma, wobei einige die Membran passieren können, andere aber nicht. Bausteine für die Herstellung von DNA und RNA gehören zu den Materialien, die in den Zellkern gelassen werden, ebenso wie Moleküle, die die Energie für den Aufbau des genetischen Materials liefern. Ribosomale Untereinheiten, die in Nukleosomen gebaut werden, sind ein Paradebeispiel für Materialien, die den Kern verlassen und ins Zytoplasma gelangen dürfen.
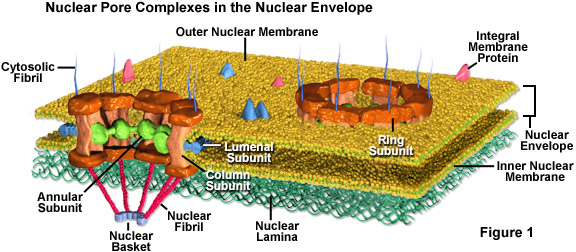
Die Kernporen sind für kleine Moleküle bis zur Größe der kleinsten Proteine vollständig durchlässig, bilden aber eine Barriere, die die meisten großen Moleküle vom Kern fernhält. Dennoch wird einigen größeren Proteinen, wie z.B. Histonen, Einlass in den Zellkern gewährt, obwohl die Poren zu klein sein müssten, um sie durchzulassen. Es wird allgemein angenommen, dass die ausgeklügelte Proteinstruktur, die als Kernporenkomplex (siehe Abbildung 1) bezeichnet wird und jede Pore umgibt, eine Schlüsselrolle dabei spielt, den aktiven Transport ausgewählter großer Moleküle in und aus dem Zellkern zu ermöglichen.
Der Kernporenkomplex besteht aus mehreren Untereinheiten. Das Innere der Pore ist von einem nichtmembranösen Material umgeben, das in einem Ring organisiert ist, der speichenartige Strukturen in Richtung des Zentrums des kleinen Kanals verlängert. Die eigentliche Porenwand besteht überwiegend aus säulenförmigen Untereinheiten, und lumenale Untereinheiten verankern mit Hilfe von Transmembranproteinen den gesamten Porenkomplex in der Kernhülle. Außerdem erstrecken sich winzige Fibrillen in der Regel von beiden Seiten des Komplexes und sammeln sich zu korbartigen Konfigurationen auf der Kernseite des Komplexes. Die proteinhaltige Zusammensetzung dieser Fibrillen ist auf den gegenüberliegenden Seiten der Struktur unterschiedlich.
Zusätzlich zu ihrer Rolle beim Kerntransport sind Kernporen als Stellen wichtig, an denen die äußere und die innere Membran der Kernhülle miteinander verschmolzen werden. Aufgrund dieser Verschmelzung können die Membranen als kontinuierlich miteinander betrachtet werden, obwohl sie unterschiedliche biochemische Eigenschaften haben und auf unterschiedliche Weise funktionieren können. Da die äußere Kernmembran auch kontinuierlich mit der Membran des endoplasmatischen Retikulums (ER) ist, können sowohl sie als auch die innere Kernmembran Membranmaterialien mit dem ER austauschen. Diese Fähigkeit ermöglicht es der Kernhülle, sich bei Bedarf zu vergrößern oder zu verkleinern, um den dynamischen Inhalt des Zellkerns aufzunehmen.
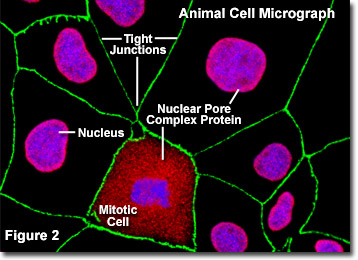
Abbildung 2 zeigt eine digitale Fluoreszenzaufnahme einer adhärenten Kultur von Madin-Darby-Hundenieren-Zellen (MDCK-Linie), die mit Fluoreszenzsonden angefärbt wurden, die auf den Zellkern zielen (blau), Kernporenkomplexproteine (rot) und die zwischen Epithelzellen gebildeten tight junctions (grün) gefärbt, um die Nähe dieser Strukturen zu demonstrieren. Die Kernporen dieser Zellen wurden mit einem polyklonalen Breitband-Antikörper gegen eine große Familie von Kernporenkomplex-Proteinen markiert, der als nützliches Werkzeug zur Untersuchung der Morphologie und Zusammensetzung des Zellkerns und der Kernhülle dient. Die Antikörpermischung ist auch nützlich, um Veränderungen in der Kernstruktur während der Mitose und Meiose zu untersuchen (beachten Sie die mitotische Zelle im unteren mittleren Teil von Abbildung 2).
Die Porendichte variiert stark und ist tendenziell am größten bei Zellen, die hoch aktiviert und differenziert sind, wie z. B. Leberzellen. Eine typische Säugetierzelle weist etwa 3.000 bis 4.000 Poren entlang ihrer Kernhülle auf. Die Eizellen bestimmter Amphibien haben jedoch so große Kerne und eine solche Dichte an Poren, dass die Kernhülle einer dieser Zellen mehr als zehn Millionen Poren enthalten kann. Folglich wurden diese Eizellen stark für Studien über Kernporenkomplexe und Kerntransport genutzt.
Zurück zum Zellkern
Zurück zur Zellstruktur von Tieren
Zurück zur Zellstruktur von Pflanzen
Fragen oder Kommentare? Schicken Sie uns eine E-Mail.
© 1995-2021 by Michael W. Davidson and The Florida State University. Alle Rechte vorbehalten. Keine Bilder, Grafiken, Software, Skripte oder Applets dürfen ohne Genehmigung der Urheberrechtsinhaber vervielfältigt oder in irgendeiner Weise verwendet werden. Mit der Nutzung dieser Website erklären Sie sich mit allen rechtlichen Bedingungen einverstanden, die von den Eigentümern festgelegt wurden.
Diese Website wird von unserem
Graphics & Web Programming Team
in Zusammenarbeit mit der Optischen Mikroskopie am
National High Magnetic Field Laboratory gepflegt.
Letzte Änderung: Freitag, 13. November 2015 um 02:18 Uhr
Zugriffszahl seit 14. Mai 2005: 157213
Mikroskope bereitgestellt von:
![]()
![]()