Wprowadzenie
Neurony nieustannie odbierają sygnały od tysięcy innych neuronów wokół nich. Jednak to, czy te dane wejściowe są w stanie wywołać potencjał czynnościowy, czy nie, zależy od sumowania tych danych. Sumowanie może być zdefiniowane jako proces, w którym sygnały pobudzające i hamujące razem są w stanie wygenerować potencjał czynnościowy lub nie.

Istnieją dwa rodzaje sumowania, są to odpowiednio sumowanie czasowe i przestrzenne. Sumowanie przestrzenne może być zdefiniowane jako sygnały pochodzące z wielu jednoczesnych wejść. Temporal summation z drugiej strony pochodzi z powtarzających się wejść. Zasadniczo, aby osiągnąć potencjał czynnościowy, musi zostać osiągnięte napięcie progowe. Można to określić przez zsumowanie poszczególnych wejść z sumowania czasowego i przestrzennego.
Są dwa rodzaje neuroprzekaźników uwalnianych z neuronów presynaptycznych. Depolaryzacja jest wywoływana przez neuroprzekaźniki pobudzające, jest to tzw. pobudzający potencjał postsynaptyczny (EPSP). Neurotransmitery hamujące powodują hiperpolaryzację, inaczej mówiąc hamujący potencjał postsynaptyczny (IPSP). Neurony są w stanie wpływać na siebie nawzajem na wiele sposobów. Są one w stanie powodować pobudzenie, hamowanie i mogą wzajemnie wpływać na swoją pobudliwość.
Zahamowanie to usunięcie pobudzających sygnałów wejściowych, które mogą ułatwić drogę. Zahamowanie z drugiej strony jest usunięciem wejścia hamującego.
Wejścia z wielu źródeł na neuron mogą być sumowane przestrzennie, jednak wejścia muszą być blisko siebie tak, aby żadne z wczesnych wejść nie zanikło. Jeśli neuron otrzymuje wiele wejść z jednego źródła w wymaganym bliskim odstępie czasu, tak że żadne z wejść nie ulega rozpadowi, to wejścia te mogą się sumować czasowo.
Innym bardzo ważnym czynnikiem decydującym o tym, czy potencjał progowy zostanie osiągnięty i powstanie potencjał czynnościowy, czy nie, jest odległość między synapsą a ciałem komórki neuronu. Im bliższa odległość między synapsą a ciałem komórki, tym większa suma i tym większe prawdopodobieństwo wywołania potencjału czynnościowego.
Jak wiemy, środkiem lokomocji dla neuronu postsynaptycznego jest dendryt. Te dendryty mają niewiele kanałów jonowych bramkowanych napięciem. Stąd, po dotarciu do ciała komórki neuronu, potencjał postsynaptyczny ulega osłabieniu. Ciało komórkowe neuronu sumuje przychodzące potencjały. Potencjał czynnościowy jest następnie wywoływany przez transmisję potencjału netto do wzgórza aksonu.
Sumowanie przestrzenne
Kiedy wejścia z wielu neuronów wywołują potencjał czynnościowy, nazywa się to sumowaniem przestrzennym. Potencjały te pochodzą najczęściej z dendrytów, dodajemy te wejścia razem, aby uzyskać sumowanie przestrzenne.
Im większa liczba pobudzających potencjałów postsynaptycznych, tym większa szansa, że potencjał osiągnie potencjał progowy do wywołania potencjału czynnościowego. Analogicznie, im większa liczba hamujących potencjałów postsynaptycznych, tym mniejsze szanse na osiągnięcie potencjału progowego do wygenerowania potencjału czynnościowego.
Szanse na wywołanie potencjału czynnościowego zależą również od tego, jak blisko wzgórza aksonalnego znajduje się wejście dendrytyczne. Im bliżej wzgórza aksonalnego znajduje się wejście dendrytyczne, tym bardziej prawdopodobne jest wywołanie potencjału czynnościowego. Shunting pobudzającego potencjału postsynaptycznego jest to zniweczenie pobudzającego wejścia przez przestrzenne zsumowanie hamujących wejść.
Sumowanie czasowe
Gdy duża ilość presynaptycznych potencjałów czynnościowych neuronu wyzwala postsynaptyczne potencjały czynnościowe, które sumują się ze sobą, nazywa się to sumowaniem czasowym.
W tym przypadku, odstęp pomiędzy potencjałami czynnościowymi jest mniejszy niż czas trwania postsynaptycznego potencjału czynnościowego. Sumowanie może być zwiększone, jeśli stała czasowa błony komórkowej jest wystarczająco długa. Kiedy zaczyna się następny potencjał postsynaptyczny, amplituda poprzedniego potencjału postsynaptycznego sumuje się z nim, wytwarzając większy potencjał zwiększający prawdopodobieństwo osiągnięcia potencjału progowego.
Mechanizm
Komórki postsynaptyczne zawierają kanały jonowe, te kanały jonowe mogą się otwierać lub zamykać w zależności od tego, który neuroprzekaźnik wiąże się z receptorami. Otwarcie/zamknięcie tych kanałów powoduje powstanie potencjału postsynaptycznego. Istnieją 2 rodzaje potencjałów postsynaptycznych. Pobudzający potencjał postsynaptyczny to taki, który zwiększa szanse na zainicjowanie potencjału czynnościowego. Podobnie hamujące potencjały postsynaptyczne zmniejszają szanse na zainicjowanie potencjału czynnościowego.
Neuroprzekaźniki pobudzające (glutaminian)
Jednym z głównych przykładów neuroprzekaźnika pobudzającego jest glutaminian. Glutaminian ten wiąże się z receptorami AMPA na błonie postsynaptycznej. Wiązanie to powoduje napływ kationu sodu. Ten napływ sodu powoduje depolaryzację. Jest to określane jako pobudzający potencjał postsynaptyczny (EPSP). Należy zauważyć, że aby suma EPSP osiągnęła potencjał progowy, konieczna jest duża liczba tych wejść. Działanie neurotransmiterów trwa znacznie dłużej niż działanie impulsów presynaptycznych.
Różnica pomiędzy pobudzającymi potencjałami postsynaptycznymi a potencjałami czynnościowymi polega na tym, że pobudzające potencjały postsynaptyczne są w stanie sumować swoje wejścia wytwarzając stopniowaną odpowiedź w przeciwieństwie do odpowiedzi typu „wszystko albo nic”, w której potencjał progowy może zostać osiągnięty stymulując potencjał czynnościowy lub może nie zostać osiągnięty w ogóle.
Inhibitory neurotransmitery (GABA)
GABA jest głównym neurotransmiterem zaangażowanym w hamujące potencjały postsynaptyczne (IPSP). Po związaniu się z receptorami neuronów postsynaptycznych GABA otwiera specyficzne kanały jonowe, które różnią się od tych otwieranych w EPSP przez neuroprzekaźniki pobudzające, takie jak glutaminian. Kanały te umożliwiają napływ ujemnie naładowanych anionów lub odpływ dodatnio naładowanych kationów. W tym przypadku anionem są jony chlorkowe. Wypływającym kationem są jony potasu. Oba jony mają ten sam efekt w obniżaniu potencjału błony, powodując hiperpolaryzację neuronu postsynaptycznego.
Sumowanie tych IPSPs i spadek napięcia błony spowoduje odchylenie od potencjału progowego hamując potencjał czynnościowy. Jednakże te IPSPs i EPSPs mogą występować w tym samym czasie, stąd neuron postsynaptyczny może otrzymywać sygnały pobudzające od glutaminianu i sygnały hamujące od GABA. Celem działania hamującego glutaminianu jest obniżenie potencjału błony z dala od potencjału progowego poprzez hiperpolaryzację.
Algebraiczne przetwarzanie EPSPs i IPSPs
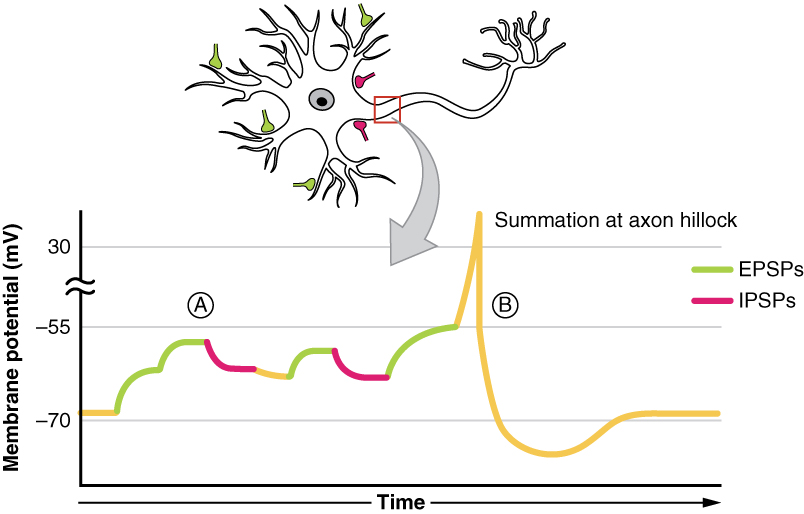
W dowolnym neuronie w dowolnym momencie, będzie on otrzymywał wiele wejść EPSP i IPSP jednocześnie. Aby określić wyjście, czy potencjał progowy zostanie osiągnięty i potencjał czynnościowy zostanie wywołany, czy nie, należy dokonać algebraicznego przetworzenia tych EPSP i IPSP. Neurony te będą otrzymywały liczne sygnały wejściowe, czy to z wielu neuronów (sumowanie przestrzenne), czy też z jednego neuronu (sumowanie czasowe).
Ta informacja wyjściowa zależy od liczby każdego typu neuroprzekaźnika, czy będą to neuroprzekaźniki pobudzające, takie jak glutaminian, które powodują napływ jonów sodu przez kanały jonowe sodu, czy też neuroprzekaźniki hamujące, takie jak GABA, które powodują napływ jonów chlorkowych lub potasu przez kanały jonowe chlorkowe lub potasowe. Ta synapsa może być określana jako punkt decyzyjny, gdzie algebraiczne przetwarzanie tych IPSPs i EPSPs określa wyjście.
Wzgórze aksonalne

Część ciała komórkowego neuronu, która łączy się z aksonem, nazywana jest wzgórzem aksonalnym. Posiada on rzadką dystrybucję substancji Nissla. Jesteśmy w stanie zidentyfikować ją za pomocą mikroskopii świetlnej. Ponieważ wzgórek aksonowy łączy akson i somatę neuronu, jest to końcowy obszar somy, w którym następuje sumowanie potencjałów błonowych z wejść synaptycznych. Ta suma jest następnie przekazywana do aksonu.
Mimo że w przeszłości wielu uważało, że wzgórek aksonu jest strefą wyzwalania, gdzie inicjowane są potencjały czynnościowe, obecnie uważa się, że początkowy odcinek pomiędzy początkowym niezmielinizowanym odcinkiem aksonu a szczytem wzgórka aksonu jest miejscem, gdzie inicjowany jest potencjał czynnościowy. Pozytywny punkt aksonu, w którym rozpoczyna się potencjał czynnościowy, różni się w zależności od komórki.
Stymulacja hormonalna i efekty drugiego posłańca neuroprzekaźnika mogą również zmienić ten pozytywny punkt. Lokalizacja potencjałów membranowych do części somalnej lub aksonalnej komórki może być osiągnięta przez wyznaczenie odrębnych domen membranowych pomiędzy aksonem a ciałem komórki. Może to być osiągnięte przez wzgórek aksonu.
Shunting
Często w neuronie, pobudzające potencjały postsynaptyczne i hamujące potencjały postsynaptyczne w dendrycie są bardzo blisko siebie. Nazywa się to shuntingiem. Oprócz dendrytów, shunting może również wystąpić w soma komórki.
Sumowanie czasowe powiedziałoby nam, aby zsumować te pobudzające i hamujące potencjały postsynaptyczne w celu określenia wyniku, czy potencjał progowy został osiągnięty i potencjał czynnościowy został zainicjowany, czy nie. Jednak w przypadku tej sekwencji zdarzeń zachodzących w soma połączenia, opór komórki jest zmieniany przez wejście hamujące. Komórka zaczyna przeciekać, to stworzy shunt, w przeciwieństwie do pozbycia się efektu wejścia pobudzającego.
Zastosowanie terapeutyczne
Przy omawianiu stymulacji nocyceptywnej, sumowanie czasowe jest powtarzalną integracją bodźców bólowych. Z kolei sumowanie przestrzenne to integracja bodźców nocyceptywnych z dużych obszarów. Wiele chorób przewlekłych ma objawy w postaci długotrwałego bólu, który jest rozległy. Dlatego też w chorobach przewlekłych stwierdza się zarówno czasowe, jak i przestrzenne sumowanie bodźców nocyceptywnych. Eksperymenty stymulacji ciśnieniowej dowodzą, że czasowa sumacja nocyceptywnych sygnałów jest ułatwiona przez sumację przestrzenną. Stąd najlepszym postępowaniem w leczeniu bólu przewlekłego jest ukierunkowanie leczenia zarówno na przestrzenną jak i czasową sumację bólu.
Badania
Większość eksperymentów nad sumacją przestrzenną jest badana na neuronach wzrokowych i czuciowych, powodem tego jest fakt, że mają one stały zakres częstotliwości neuronów pobudzających i hamujących. Tłumienie potencjałów postsynaptycznych w ciałach komórkowych neuronów i dendrytach jest w ostatnich czasach głównym tematem badań nad sumowaniem neuronalnym. Ze względu na fakt, że odpowiedź tych interakcji jest mniejsza niż suma indywidualnych odpowiedzi, odpowiedzi te są określane jako nieliniowe. Shunting jest częstą przyczyną tego efektu. Shunting to zmniejszone przewodnictwo pobudzającego potencjału postsynaptycznego.
Naoki Kogo i Michael Ariel byli w stanie przeprowadzić eksperyment na jądrze podstawy nerwu wzrokowego żółwia, z którego udało im się wyciągnąć informacje na temat hamowania przez shunning. Według ich pracy, sumowanie przestrzenne EPSP i IPSP powodowało tłumienie odpowiedzi pobudzającej podczas odpowiedzi hamującej. Zauważono również, że po tłumieniu następowała augmentacja odpowiedzi pobudzającej. Kontrolą dla tego eksperymentu było badanie tłumienia, gdy prąd hiperpolaryzacji aktywował te wrażliwe na napięcie kanały. Stwierdzono, że tłumienie jest spowodowane przez otwarcie kanału receptora synaptycznego, a nie hiperpolaryzację.
Wniosek
W każdej chwili neuron otrzymuje wiele danych wejściowych z wielu różnych neuronów.
Te wejścia mogą być pobudzające lub hamujące, jednak aby określić, czy potencjał progowy zostanie osiągnięty i potencjał czynnościowy zostanie wywołany, czy nie, należy wziąć pod uwagę wiele czynników.
Różne neuroprzekaźniki mają różne efekty, pobudzające neuroprzekaźniki jak glutaminian powodują otwarcie kanałów jonów sodu powodując napływ jonów sodu podnosząc potencjał błony. Neurotransmitery hamujące, takie jak GABA, mogą otwierać kanały jonów chlorkowych, powodując napływ jonów chlorkowych, lub mogą otwierać kanały jonów potasowych, powodując odpływ jonów potasowych.
Oba te efekty obniżają potencjał błonowy, odchylając go od napięcia progowego, co zmniejsza szanse na zainicjowanie potencjału czynnościowego.
Innym czynnikiem zwiększającym szanse na zainicjowanie potencjału czynnościowego jest bliskość wejścia dendrytycznego do wzgórka aksonalnego, im bliżej tym większe szanse na zainicjowanie potencjału czynnościowego. Wzgórek aksonowy jest częścią somy neuronu, która łączy go z aksonem.
Neuroprzekaźniki pobudzające zwiększają szanse na depolaryzację, zmiana w napięciu błony jest nazywana pobudzającym potencjałem postsynaptycznym (EPSP). Neuroprzekaźniki hamujące zwiększają szanse na hiperpolaryzację, ta zmiana w potencjale błonowym nazywana jest hamującym potencjałem postsynaptycznym (IPSP).
To, czy napięcie progowe zostanie osiągnięte i potencjał czynnościowy zostanie zainicjowany czy nie, zależy od algebraicznego sumowania poszczególnych wejść. Istnieją dwa rodzaje sumowania.
Sumowanie wielu wejść z wielu różnych neuronów presynaptycznych wyzwalających potencjał czynnościowy nazywane jest sumowaniem przestrzennym. Wysoka częstotliwość wejść z jednego neuronu presynaptycznego zsumowanych w celu wytworzenia potencjału czynnościowego nazywana jest sumowaniem czasowym. Ten czas trwania potencjału postsynaptycznego jest dłuższy niż czas trwania pomiędzy odpowiednimi potencjałami czynnościowymi.