Trawienie i wchłanianie lipidów to złożone procesy. Biorą w nich udział rozpuszczalne enzymy, substraty o różnym stopniu rozpuszczalności, a zachodzą głównie w żołądku i jelicie cienkim.
Dietetyczne lipidy to trójglicerydy, fosfolipidy, steroidy, zwłaszcza cholesterol i estry cholesterolu, witaminy rozpuszczalne w tłuszczach, czyli witamina A, D, E i K oraz karotenoidy.
Lipidy mogą mieć postać stałą lub ciekłą w temperaturze pokojowej i są określane odpowiednio jako tłuszcze i oleje.
- Trójglicerydy lub triacyloglicerole stanowią około 90% lipidów pokarmowych. Składają się one z jednej cząsteczki glicerolu zestryfikowanej z trzema kwasami tłuszczowymi, głównie długołańcuchowymi (16-20 atomów węgla). Mają ponad dwukrotnie większą gęstość energetyczną niż węglowodany (9 kcal/g vs. ok. 3,75) i aminokwasy. Muszą one uwalniać swoje kwasy tłuszczowe, aby mogły być wykorzystane jako źródło energii.
- Fosfolipidy, główne składniki błon biologicznych, składają się z jednej cząsteczki glicerolu zestryfikowanej z dwoma kwasami tłuszczowymi w pozycjach sn-1 i sn-2 oraz kwasem fosforowym w pozycji sn-3. Z kolei grupa fosforanowa łączy się z grupą hydrofilową, taką jak cholina, seryna lub inozytol, poprzez wiązanie estrowe. Dzienne spożycie fosfolipidów jest niskie, 1-2 g, jednak również fosfolipidy żółciowe wlewają się do jelita cienkiego, około 10-20 g dziennie, głównie fosfatydylocholina.
- Cholesterol i jego estry, wraz z niewielkimi ilościami hormonów steroidowych, występują tylko w produktach zwierzęcych, w przeciwieństwie do lipidów widzianych do tej pory, które występują również w produktach roślinnych.
W jelicie cienkim, oprócz cholesterolu pokarmowego (który nie powinien przekraczać 300 mg/dobę), znajduje się również cholesterol żółciowy, około 1 g/dobę. Zarówno cholesterol pokarmowy, jak i żółciowy występują głównie w postaci niezestryfikowanej, około 85-90%, jedynej formy cholesterolu, która może być wchłonięta w jelicie cienkim.
Podobnie jak witaminy, nawet ta cząsteczka steroidowa nie jest źródłem energii.
Różne stanole i sterole roślinne, w szczególności β-sitosterol (który nie jest wchłaniany w warunkach fizjologicznych), są również zaliczane do steroidów dietetycznych.
Mimo że towarzystwa naukowe zalecają spożycie lipidów (głównie triacylogliceroli) nieprzekraczające 30% dziennego spożycia kalorii, w diecie zachodniej tłuszcze i oleje dostarczają od 30 do 45% dziennego spożycia kalorii.
Hydrofobowość, jedna z charakterystycznych właściwości wielu dietetycznych lipidów, która sprawia, że trójglicerydy są doskonałymi cząsteczkami do magazynowania energii, stwarza problemy, gdy takie cząsteczki są trawione w przewodzie pokarmowym, wchłaniane w jelicie cienkim i wreszcie transportowane w krążeniu po wchłonięciu lub mobilizacji z zapasów ciała.
Indeed, lipidy, takie jak trójglicerydy z długim łańcuchem kwasów tłuszczowych, a cholesterol i rozpuszczalne w tłuszczach estry witamin są niezwykle hydrofobowe, i agregacji w duże kropelki w żołądku i jelicie cienkim. Te kropelki są następnie emulgowane, aby umożliwić hydrolazom katalizowanie trawienia lipidów.
KONKURS
- Kroki, enzymy i produkty
- Trawienie lipidów i lipaza językowa
- Trawienie lipidów i lipaza żołądkowa
- Sole żółciowe i emulsja kropli lipidów
- Trawienie lipidów i lipaza trzustkowa
- Trawienie lipidów i lipaza w mleku kobiecym
- Trawienie lipidów i esteraza cholesterolowa
- Trawienie lipidów i fosfolipazy
Stepy, enzymy i produkty
Trawienie lipidów rozpoczyna się w jamie ustnej, kontynuowane jest w żołądku, a kończy w jelicie cienkim.nzymy biorące udział w trawieniu triacylogliceroli nazywane są lipazami (EC 3.1.1.3). Są to białka, które katalizują częściową hydrolizę trójglicerydów do mieszaniny wolnych kwasów tłuszczowych i acylogliceroli. Istnieje kilka lipaz, z których najważniejsza jest produkowana przez zewnątrzwydzielniczą trzustkę; inne to lipaza językowa, lipaza żołądkowa i lipaza mleka kobiecego.

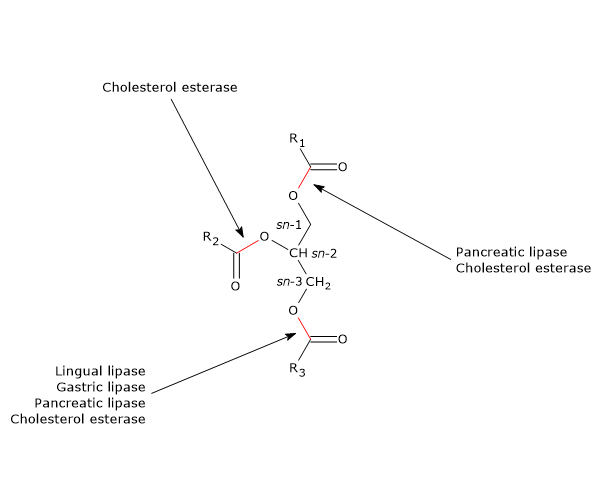
Innymi enzymami biorącymi udział w trawieniu lipidów są esteraza cholesterolowa oraz fosfolipazy A1 i A2.
Trawienie lipidów i lipaza językowa
W jamie ustnej pokarm jest rozbijany na małe cząstki i mieszany z lipazą językową.
Ezym jest produkowany i wydzielany przez surowicze gruczoły językowe, zwane również gruczołami von Ebnera.est stabilny w środowisku kwaśnym, dlatego pozostaje aktywny w żołądku, a także w jelicie cienkim w przypadku, gdy nie ma prawidłowego wydzielania wodorowęglanów przez trzustkę.
Reakcja katalizowana przez enzym uwalnia pojedynczy kwas tłuszczowy, najlepiej krótkołańcuchowy lub średniołańcuchowy, oraz 1,2-diacyloglicerol, który jest następnie hydrolizowany w dwunastnicy.
Uwaga: krótkołańcuchowe kwasy tłuszczowe są głównie estryfikowane w pozycji sn-3 triacyloglicerolu.
Lingwalna lipaza odgrywa skromną rolę w trawieniu triacylogliceroli, ponieważ:
- ma wolniejszą aktywność niż lipaza trzustkowa;
- działa w żołądku, środowisku wodnym, w którym lipidy mają tendencję do koalescencji, tworząc oddzielną fazę od otaczającego środowiska, ograniczając w ten sposób możliwości enzymu do hydrolizy triacylogliceroli.
Aktywność lipazy językowej nie jest szczególnie ważna dla zdrowych dorosłych. Wręcz przeciwnie, jest bardzo ważna dla niemowląt, u których lipaza trzustkowa jest jeszcze niedojrzała, również uprzywilejowana przez fakt, że trójglicerydy mleka są bogate w krótkołańcuchowe i średniołańcuchowe kwasy tłuszczowe. Ponadto, podobnie jak lipaza żołądkowa (patrz poniżej), jest w stanie przeniknąć do kuleczek tłuszczu w mleku, inicjując w ten sposób proces trawienia (lipaza trzustkowa nie jest w stanie przeniknąć do tych kuleczek tłuszczu).
Oczywiście, działanie tego enzymu jest ważne również wtedy, gdy lipaza trzustkowa jest nieobecna.
Jako że język jest wrażliwy na smak wolnych kwasów tłuszczowych, zwłaszcza wielonienasyconych, a nie trójglicerydów, aktywność lipazy językowej może odgrywać rolę w wykrywaniu tłustych pokarmów jako źródła energii, a tym samym wpływać na wybory żywieniowe.
Wreszcie, uwalnianie krótkołańcuchowych i średniołańcuchowych kwasów tłuszczowych i diacylogliceroli jest ważne również dlatego, że są to cząsteczki amfipatyczne, to znaczy, że mają region hydrofilowy, który oddziałuje z otaczającą fazą wodną, oraz region hydrofobowy, który jest zorientowany w kierunku rdzenia kropli lipidowych. Ze względu na działanie tych środków powierzchniowo czynnych, kropelki tłuszczu uzyskują powierzchnię hydrofilową, tj. stabilny interfejs z otaczającą fazą wodną. To, wraz z działaniem burczenia żołądka, prowadzi do powstania emulsji złożonej z kropel, które zmniejszają swój rozmiar. Emulsja jest następnie uwalniana do dwunastnicy w postaci śluzu. Inne amfipatyczne cząsteczki obecne w żywności to lecytyna i fosfolipidy, a wszystkie razem pozwalają na zwiększenie powierzchni dostępnej dla aktywności hydrolazy.
W żołądku lipidy mieszają się również z sokiem żołądkowym i kwasem solnym. Kwas ten denaturuje również białka kompleksów lipidowo-białkowych; polipeptydy są następnie trawione przez proteazy żołądkowe, a lipidy są uwalniane.
Trawienie lipidów i lipaza żołądkowa
W żołądku lipidy są hydrolizowane również przez drugą kwaśną lipazę – lipazę żołądkową. Enzym ten jest wydzielany przez główne komórki błony śluzowej żołądka i ma optymalne pH około 4, ale jest nadal dość aktywny przy mniej kwaśnych wartościach pH, 6 do 6,5. Dlatego prawdopodobnie pozostaje aktywny nawet w górnej części dwunastnicy, gdzie pH wynosi od 6 do 7.
Ezym preferencyjnie katalizuje hydrolizę trójglicerydów z krótkołańcuchowymi i średniołańcuchowymi kwasami tłuszczowymi, ale może również hydrolizować długołańcuchowe kwasy tłuszczowe. Niezależnie od rodzaju kwasów tłuszczowych, lipaza żołądkowa preferencyjnie rozszczepia te w pozycji sn-3, prowadząc do uwolnienia wolnego kwasu tłuszczowego i 1,2-diacyloglicerolu, cząsteczek, które mogą działać jako środki powierzchniowo czynne, jak wcześniej zauważono.
Podobnie jak lipaza językowa, jest ona szczególnie aktywna w stosunku do trójglicerydów mleka, również mleka matki, które są bogate w krótkołańcuchowe i średniołańcuchowe kwasy tłuszczowe. Ponadto jest również aktywna w stosunku do triacylogliceroli niektórych olejów tropikalnych, np. oleju kokosowego, które są bogate w średniołańcuchowe kwasy tłuszczowe.
Ezym ten może odpowiadać za 10-30% hydrolizy triacylogliceroli zachodzącej w przewodzie pokarmowym, a do 50% u niemowląt karmionych piersią.
Sole żółciowe i emulsja kropli lipidowych
Płyn żółciowy, zawierający emulsję lipidową złożoną z kropli o średnicy mniejszej niż 0,5 mm, dostaje się do górnej części jelita cienkiego, dwunastnicy, gdzie kontynuowana jest hydroliza triglicerydów.

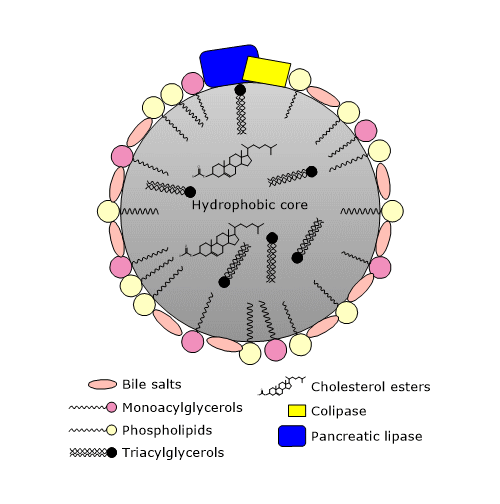
W dwunastnicy, żółć jest mieszana z żółcią, której uwalnianie przez pęcherzyk żółciowy jest stymulowane przez cholecystokininę, hormon wydzielany przez komórki błony śluzowej dwunastnicy i jelita czczego w odpowiedzi na spożycie posiłku, zwłaszcza wysokotłuszczowego. W żółci, wśród innych składników, znajdują się sole żółciowe, fosfolipidy i cholesterol. Sole żółciowe to kwasy żółciowe sprzężone z glicyną lub tauryną. Z kolei kwasy żółciowe są utlenionymi pochodnymi cholesterolu. Zarówno kwasy żółciowe, jak i sole żółciowe są syntetyzowane przez wątrobę. Są cząsteczkami amfipatycznymi, w których planarnej strukturze pierścienia można wyróżnić powierzchnię hydrofobową i hydrofilową. Dlatego są one w stanie dodatkowo emulgować kropelki lipidów, zwiększając powierzchnię dla aktywności hydrolaz.
W szczególności sole kwasu cholowego, które zawierają trzy grupy hydroksylowe, są lepszymi emulgatorami niż sole kwasu deoksycholowego, które zawierają tylko dwie grupy hydroksylowe.
Uwaga: pęcherzyk żółciowy wydziela około 30 g soli żółciowych każdego dnia, wraz z fosfolipidami i cholesterolem. Większość soli żółciowych i cholesterolu jest następnie wchłaniany ponownie, tak, że dzienna utrata soli żółciowych i steroidów w kale jest dość niska, 0.2-1 g.
Mechanizm perystaltyki i surfaktantów poznanych do tej pory (wolne kwasy tłuszczowe, acyloglicerole, fosfolipidy i sole żółciowe) zapewniają tworzenie mikroskopijnych miceli, które dodatkowo zwiększają dostępną powierzchnię dla aktywności enzymów hydrolitycznych.
Należy podkreślić, że triacyloglicerole z krótkołańcuchowymi i średniołańcuchowymi kwasami tłuszczowymi mogą być zarówno hydrolizowane, jak i wchłaniane przy braku soli żółciowych, chociaż ich obecność zwiększa wchłanianie.
Trawienie lipidów i lipaza trzustkowa
Cholecystokinina stymuluje również zewnątrzwydzielniczą trzustkę do wydzielania soku trzustkowego zawierającego m.in. lipazę trzustkową. Enzym ten katalizuje trawienie większości spożytych trójglicerydów, głównie w górnej części jelita czczego, a jego optimum pH wynosi 7,0 do 8,8 (nie jest więc lipazą kwaśną, jak lipazy językowe i żołądkowe). Katalizuje ona rozszczepianie kwasów tłuszczowych, zwykle o 10 lub więcej atomach węgla, głównie w pozycjach sn-1 i sn-3 szkieletu glicerolu. Produktami reakcji są wolne kwasy tłuszczowe i 2-monoacyloglicerole. 2-monoacyloglicerol, główna postać, w której monoacyloglicerole są wchłaniane z jelita cienkiego, może ulec procesowi izomeryzacji, w którym reszta kwasu tłuszczowego przechodzi na węgiel 1 lub 3. Jednakże tempo izomeryzacji jest wolniejsze niż tempo wchłaniania cząsteczki z jelita cienkiego. W warunkach in vitro lipaza trzustkowa jest hamowana przez sole żółci, natomiast in vivo hydrolizuje trójglicerydy w sposób bardzo wydajny, dzięki obecności kofaktora białkowego wydzielanego przez zewnątrzwydzielniczą część trzustki – kolipazy. Białko to nie ma aktywności katalitycznej, jest produkowane w postaci nieaktywnej, zwanej prokolipazą, a aktywowane jest przez trypsynę w dwunastnicy. Kropelki lipidowe są pokryte fosfolipidami i solami żółci, które nadają im ładunek ujemny, uniemożliwiający wiązanie lipazy, ale przyciągający kolipazę. Z kolei kolipaza wiąże lipazę trzustkową (lipaza i kolipaza wiążą się w stosunku molowym 1:1), zakotwiczając w ten sposób enzym na wodno-lipidowej powierzchni kropli lipidowych.
Trawienie lipidów i lipaza w mleku matki
Lipaza ta ma neutralne optimum pH i jest stymulowana przez sole żółci. Enzym ten w znacznym stopniu przyczynia się do hydrolizy trójglicerydów w jelitach niemowląt karmionych piersią.
W mleku krowim natomiast lipazy nie występują.
Trawienie lipidów a esteraza cholesterolowa
Kolejnym enzymem obecnym w soku trzustkowym jest esteraza cholesterolowa (EC 3.1.1.13). Enzym ten, syntetyzowany i wydzielany w aktywnej postaci przez zewnątrzwydzielniczą część trzustki, jest lipazą o szerokiej specyficzności, aktywną wobec:
- estry cholesterolu, z cholesterolem i wolnymi kwasami tłuszczowymi jako produktami reakcji;
- triglicerydów, w których hydrolizuje wszystkie trzy wiązania estrowe, z tego powodu nazywana jest również esterazą niespecyficzną (hydrolizuje 10%-15% triglicerydów pokarmowych);
- monoacylogliceroli;
- fosfolipidów;
- estrów witamin A i D.

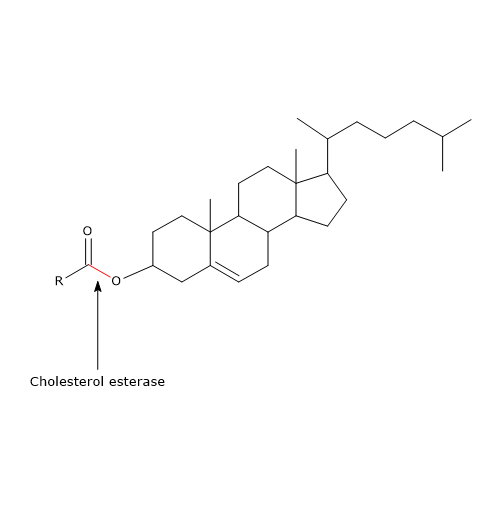
Podobnie jak fosfolipaza A2 (patrz poniżej), jest ona aktywna głównie w stosunku do estrów cholesterolu wbudowanych w micele soli żółciowych. W przeciwieństwie do lipazy trzustkowej, jej aktywność jest stymulowana przez sole żółci, głównie sole trójhydroksylowe, takie jak taurocholan sodu i glikocholan. Sole te indukują zmianę konformacyjną białka, która aktywuje enzym. Ponadto, sole trójhydroksylowe promują jego samoasocjację w polimeryczne agregaty, które chronią go przed działaniem proteaz w świetle jelita.
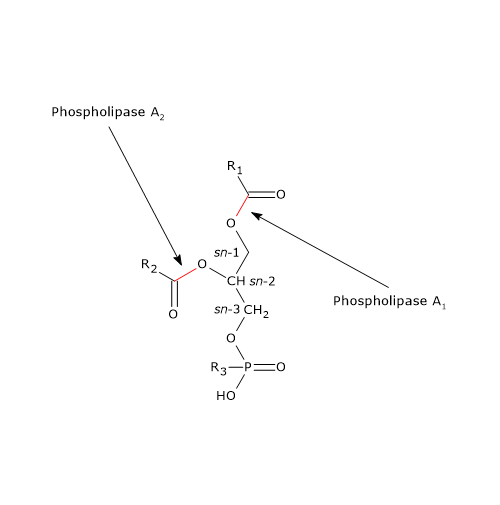
Trawienie lipidów i fosfolipazy
Trawienie fosfolipidów jest przeprowadzane przez fosfolipazy, głównie fosfolipazę A2 (EC 3.1.1.4). Enzym ten występuje w soku trzustkowym w postaci zymogenu, zwanego profosfolipazą A2, i jest aktywowany przez trypsynę, enzym biorący udział w trawieniu białek. Fosfolipaza A2 katalizuje specyficznie rozszczepienie kwasu tłuszczowego w pozycji sn-2 fosfolipidów, przy czym wykazuje szeroką specyficzność zarówno w odniesieniu do długości łańcucha węglowego docelowego kwasu tłuszczowego, jak i polarnych grup głównych fosfolipidów.
Jak już wcześniej zauważono, większość fosfolipidów w świetle jelita jest pochodzenia żółciowego, a tylko niewielka ich część pochodzi z diety. W żółci fosfolipidy tworzą micele z cholesterolem i solami żółci, a w świetle jelita są one rozmieszczone między kropelkami lipidów i tych miceli, z preferencją dla tych ostatnich. A w micelach fosfolipidy, głównie fosfatydylocholina, pełnią rolę substratu. W przypadku fosfatydylocholiny produktami reakcji są wolny kwas tłuszczowy i lizofosfatydylocholina (lizofosfolipid).
W soku trzustkowym obecna jest również fosfolipaza A1, która usuwa kwas tłuszczowy w pozycji sn-1 fosfolipidu.
W błonie śluzowej jelit wydaje się istnieć trzecia, skromna, aktywność fosfolipazy, dzięki wewnętrznemu enzymowi błonowemu. Enzym ten nazywany jest fosfolipazą B lub hydrolazą estrów retinylu, aktywną również na estrach witaminy A.
Trawienie fosfolipidów może zakończyć się powstaniem wolnego kwasu tłuszczowego i lizofosfolipidu lub może być całkowite.
Bender D.A. Wprowadzenie do żywienia i metabolizmu. 3rd Edition. Taylor & Francis, 2004
Berdanier C.D., Dwyer J., Feldman E.B. Podręcznik żywienia i żywności. 2th Edition. CRC Press. Taylor & Francis Group, 2007
Chow Ching K. „Kwasy tłuszczowe w żywności i ich implikacje zdrowotne” 3th Edition. CRC Press Taylor & Francis Group, 2008
Hamosh M. Lingual and gastric lipases. Nutrition 1990;6(6):421-8.
Iqbal J. and Hussain M.M. Intestinal lipid absorption. Am J Physiol Endocrinol Metab 2009;296:E1183-94. doi:10.1152/ajpendo.90899.2008
Nelson D.L., Cox M.M. Lehninger. Principles of biochemistry. 4th Edition. W.H. Freeman and Company, 2004
Rosenthal M.D., Glew R.H. Medical Biochemistry – Human Metabolism in Health and Disease. John Wiley J. & Sons, Inc, Publication, 2009
Stipanuk M.H., Caudill M.A. Biochemiczne, fizjologiczne i molekularne aspekty żywienia człowieka. 3rd Edition. Elsevier health sciences, 2013