Introdução
Neurões estão continuamente a receber de milhares de outros neurónios à sua volta. No entanto, se estes inputs são capazes de desencadear um potencial de acção ou não, depende da soma destes inputs. A soma pode ser definida como um processo pelo qual os sinais excitatórios e inibitórios juntos são capazes de gerar ou não um potencial de acção.

Existem dois tipos de totalizações, estas são as totalizações temporal e espacial, respectivamente. As somações espaciais podem ser definidas como sinais provenientes de múltiplas entradas simultâneas. A soma temporal, por outro lado, provém de entradas repetidas. Essencialmente para atingir um potencial de acção, a tensão limite deve ser atingida. Isto pode ser determinado pela soma das entradas individuais da soma temporal e espacial.
Existem dois tipos de neurotransmissores libertados de neurónios pré-sinápticos. A despolarização é causada por neurotransmissores excitatórios, a que se chama potencial pós-sináptico excitatório (EPSP). Os neurotransmissores inibidores causam hiperpolarização, por outras palavras, um potencial pós-sináptico inibitório (IPSP). Os neurónios são capazes de se afectar uns aos outros de várias maneiras. São capazes de causar excitação, inibição e podem enviesar a excitabilidade um do outro.
Defacilitação é a remoção de inputs excitatórios que podem facilitar uma via. A desinibição, por outro lado, é a remoção de entradas inibitórias.
Múltiplas entradas de fonte num neurónio podem ser somadas espacialmente, contudo as entradas devem ser espaçadas de modo a que nenhuma das entradas iniciais se decomponha. Se um neurónio estiver a receber múltiplas entradas de uma única fonte no intervalo de tempo próximo necessário para que nenhuma entrada se decomponha, então estas entradas podem ser somadas temporariamente.
Outro factor muito importante para determinar se o potencial limiar é alcançado e se um potencial de acção é alcançado ou não é a distância entre a sinapse e o corpo da célula neuronal. Quanto mais próxima for a distância entre a sinapse e o corpo celular, maior é a soma e maior é a probabilidade de se obter um potencial de acção.
Como sabemos, o meio de viagem de um neurónio pós-sináptico é um dendrito. Estes dendritos têm poucos canais de iões fechados de voltagem. Assim, ao atingir o corpo celular neuronal, o potencial pós-sináptico atenua-se. Este corpo de célula neuronal soma estes potenciais de entrada. Um potencial de acção é então desencadeado na transmissão do potencial líquido para o axon hillock.
Soma Espacial
Quando as entradas de múltiplos neurónios desencadeiam um potencial de acção, este é chamado como soma espacial. Estes potenciais são mais frequentemente provenientes de dendritos, adicionamos estes inputs em conjunto para obter a soma espacial.
Quanto maior for o número de potenciais excitatórios pós-sinápticos, maiores são as probabilidades do potencial atingir o potencial limiar para desencadear um potencial de acção. Da mesma forma, quanto maior for o número de potenciais pós-sinápticos inibitórios, menores as hipóteses de atingir o potencial limiar para gerar um potencial de acção.
As probabilidades de eliciar um potencial de acção são também significativamente influenciadas pelo quão próximo está o input dendrítico do axon hillock. Quanto mais próximo o input dendrítico estiver do axon hillock, maior é a probabilidade de causar um potencial de acção. O desvio de um potencial pós-sináptico excitatório é a anulação de um input excitatório pela soma espacial de inputs inibitórios.
Soma Temporal
Quando uma grande quantidade de potenciais de acção pré-sinápticos de neurónios desencadeia potenciais de acção pós-sinápticos que somam uns com os outros, isto é chamado como soma temporal.
Neste caso, o intervalo entre os potenciais de acção é inferior à duração do potencial de acção pós-sináptica. O somatório pode ser aumentado se a constante de tempo da membrana celular for suficientemente longa. Quando o potencial pós-sináptico seguinte começa, a amplitude do potencial pós-sináptico anterior irá somar-se com ele produzindo um potencial maior aumentando a probabilidade de atingir o potencial limiar.
Mecanismo
As células pós-sinápticas contêm canais iónicos, estes canais iónicos podem abrir-se ou fechar-se dependendo do neurotransmissor que se liga aos receptores. A abertura/fechamento destes canais cria um potencial pós-sináptico. Existem 2 tipos de potenciais pós-sinápticos. Um potencial pós-sináptico excitatório é aquele que aumenta as hipóteses de iniciar um potencial de acção. Do mesmo modo, os potenciais pós-sinápticos inibidores diminuem as hipóteses de iniciar um potencial de acção.
Neurotransmissores excitatórios (Glutamato)
Um exemplo principal de um neurotransmissor excitatório é o glutamato. Este glutamato liga-se aos receptores da AMPA na membrana pós-sináptica. Esta ligação provoca um influxo de catiões de sódio. Este influxo de sódio causa uma despolarização. Isto é chamado de potencial pós-sináptico excitatório (EPSP). É essencial notar que para que a soma do EPSP atinja o potencial limiar, é necessário um grande número destes inputs. Os efeitos dos neurotransmissores duram muito mais tempo do que os dos impulsos pré-sinápticos.
A diferença entre os potenciais pós-sinápticos excitatórios e os potenciais de acção é que os potenciais pós-sinápticos excitatórios são capazes de somar as suas entradas produzindo uma resposta graduada em oposição à resposta tudo ou nada em que o potencial limiar pode ser alcançado estimulando um potencial de acção ou pode não ser alcançado de todo.
Neurotransmissores inibitórios (GABA)
GABA é o principal neurotransmissor envolvido nos potenciais pós-sinápticos inibitórios (IPSP). Na ligação a receptores neuronais pós-sinápticos, GABA abre canais iónicos específicos que são diferentes dos abertos em EPSP por neurotransmissores excitatórios como o Glutamato. Estes canais permitem um influxo de ânions com carga negativa ou um efluxo de catiões com carga positiva. O ânion, neste caso, são iões cloreto. Os efluxos de catiões são iões de potássio. Ambos os iões têm o mesmo efeito na diminuição do potencial de membrana causando hiperpolarização do neurónio pós-sináptico.
Sumação destes IPSPs e a queda da tensão da membrana desviar-se-ão do potencial limiar inibidor de um potencial de acção. Contudo, estes IPSPs e EPSPs podem estar a ocorrer ao mesmo tempo, daí que o neurónio pós-sináptico possa estar a receber sinais excitatórios do glutamato e sinais inibitórios do GABA. O objectivo do glutamato inibitório é de baixar o potencial de membrana para longe do potencial limiar através da hiperpolarização.
Processamento algébrico de EPSPs e IPSPs
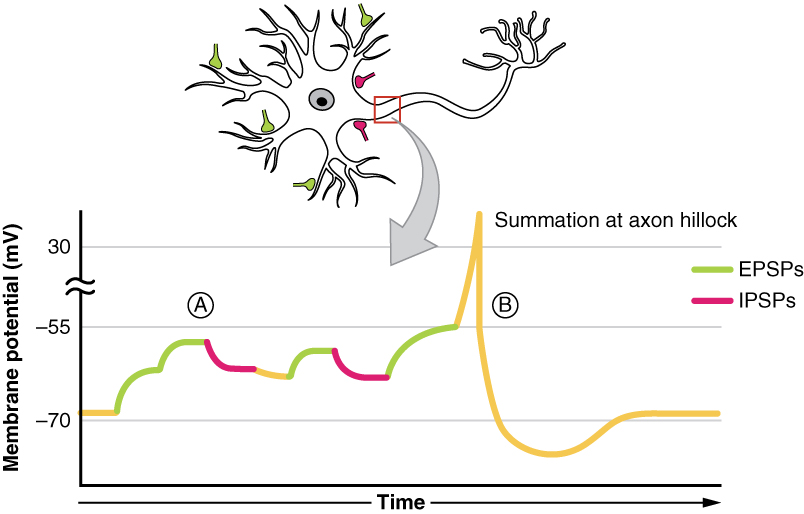
Em qualquer neurónio, em qualquer momento, receberá simultaneamente numerosas entradas de EPSP e IPSP. Para determinar se o potencial limiar será atingido e se será ou não desencadeado um potencial de acção, o processamento algébrico destes EPSPs e IPSPs deve ser tomado. Estes neurónios receberão numerosas entradas quer sejam de múltiplos neurónios (soma espacial) ou múltiplas entradas de um único neurónio (soma temporal).
Esta saída depende do número de cada tipo de neurotransmissor, quer sejam neurotransmissores excitatórios como o glutamato que causam o influxo de iões de sódio através de canais de iões de sódio ou neurotransmissores inibidores como GABA que causam um influxo de iões cloreto ou de iões de potássio através de canais de iões cloreto ou de potássio, respectivamente. Esta sinapse pode ser referida como o ponto de decisão onde o processamento algébrico destes IPSPs e EPSPs determina a saída.
Axon Hillock

A parte do corpo celular de um neurónio que se liga ao axónio é chamada axon hillock. Tem uma distribuição esparsa da substância Nissl. Somos capazes de o identificar por microscopia ligeira. Como este axon hillock liga o axónio e o soma do neurónio, é a região final do soma onde ocorre a soma dos potenciais de membrana a partir das entradas sinápticas. Este somatório é então transmitido ao axônio.
Embora no passado, muitos acreditavam que o axon hillock era a zona de desencadeamento onde os potenciais de acção eram iniciados, acredita-se agora que o segmento inicial entre o segmento axon não mielinizado inicial e o pico do axon hillock é onde o potencial de acção é iniciado. O ponto positivo do axônio onde o potencial de acção é iniciado varia de célula para célula.
Estimulação hormonal e efeitos neurotransmissores do segundo mensageiro também podem alterar este ponto positivo. A localização dos potenciais de membrana para a parte somal ou axonal da célula pode ser conseguida através da delimitação de domínios de membrana distintos entre o axónio e o corpo celular. Isto pode ser conseguido pelo axon hillock.
Shunting
Muitas vezes num neurónio, os potenciais pós-sinápticos excitatórios e inibitórios pós-sinápticos num dendrito estão muito próximos uns dos outros. A isto chama-se “shunting” (manobras). Para além dos dendritos, as manobras também podem ocorrer no somatório de uma célula.
Soma temporal dir-nos-ia para somar estes potenciais pós-sinápticos excitatórios e inibitórios pós-sinápticos, a fim de determinar o produto resultante, quer o potencial limiar seja atingido e um potencial de acção seja iniciado ou não. Contudo, no caso desta sequência de eventos que ocorrem no somatório de uma chamada, a resistência da célula é alterada pela entrada inibitória. A célula começa a vazar, o que cria um shunt em vez de se livrar do efeito excitatório do input.
Aplicação terapêutica
Quando se discute a estimulação nociceptiva, o somatório temporal é a integração repetitiva de estímulos dolorosos. O somatório espacial, por outro lado, é a integração do input nociceptivo de grandes áreas. Muitas doenças crónicas têm sintomas de longa duração de dor que são generalizados. Assim, tanto a soma temporal como a espacial nociceptiva são encontradas nas doenças crónicas. As experiências de estimulação por pressão fornecem provas de que a soma temporal dos insumos nociceptivos é facilitada através da soma espacial. Assim, a melhor linha de acção no tratamento da dor crónica é direccionar o tratamento tanto para a soma espacial como temporal da dor.
Pesquisa
A maior parte das experiências de soma espacial são testadas em neurónios ópticos e sensoriais, sendo a razão para tal o facto de terem uma gama constante de frequência de neurónios excitatórios e inibitórios. A atenuação do potencial pós-sináptico dos corpos celulares e dendritos dos neurónios é um grande foco da soma neural em estudos recentes. Devido ao facto de a resposta destas interacções ser menor que a soma das respostas individuais, estas respostas são descritas como não lineares. O shunting é uma causa comum deste efeito. O shunting é uma diminuição da condutância potencial excitatória pós-sináptica.
Naoki Kogo e Michael Ariel foram capazes de experimentar o núcleo óptico basal da tartaruga, a partir do qual foram capazes de retirar informações sobre a inibição de manobras. De acordo com o seu trabalho, a soma espacial EPSP e IPSP causou atenuação da resposta excitatória durante a resposta inibitória. Notou-se também que após a atenuação houve um aumento da resposta excitatória. O controlo para esta experiência estava a testar a atenuação quando uma corrente de hiperpolarização activou estes canais sensíveis à tensão. Concluiu-se que a atenuação é causada pela abertura do canal receptor sináptico e não pela hiperpolarização.
Conclusão
A qualquer momento, um neurónio receberá múltiplas entradas de múltiplos neurónios diferentes.
Estes inputs podem ser excitatórios ou inibitórios, contudo, para determinar se o potencial limiar será atingido e se será ou não desencadeado um potencial de acção, há uma série de factores que devem ser analisados.
Diferentes neurotransmissores têm efeitos diferentes, neurotransmissores excitatórios como o glutamato provocam a abertura de canais de iões de sódio que provocam o influxo de iões de sódio elevando o potencial de membrana. Os neurotransmissores inibidores como o GABA podem abrir canais de iões cloreto causando o influxo de iões cloreto ou podem abrir canais de iões potássio causando efluxo de iões potássio.
Bambos destes efeitos diminuem o potencial da membrana desviando-a da tensão limite, diminuindo as hipóteses de iniciar um potencial de acção.
Outro factor que aumenta as probabilidades de iniciar um potencial de acção é o quão perto está a entrada dendrítica do axon hillock, quanto mais perto, maiores as probabilidades de iniciar um potencial de acção. Um axon hillock é parte do soma de um neurónio que o liga ao axon.
Os neurotransmissores excitatórios aumentam as hipóteses de despolarização, a alteração da tensão da membrana é chamada de potencial pós-sináptico excitatório (EPSP). Os neurotransmissores inibidores aumentam as probabilidades de hiperpolarização, esta alteração no potencial de membrana chamada como potencial pós-sináptico inibitório (IPSP).
Se a tensão limite será atingida, e um potencial de acção será iniciado ou não, depende da soma algébrica das entradas individuais. Há dois tipos de somatória.
Soma de múltiplas entradas de múltiplos neurónios pré-sinápticos diferentes, desencadeando um potencial de acção, é chamada de soma espacial. Uma alta frequência de entradas de um neurónio pré-sináptico somado para produzir um potencial de acção é chamada como soma temporal. Esta duração do potencial pós-sináptico é maior do que a duração entre os respectivos potenciais de acção.