Einführung
Neuronen erhalten kontinuierlich von tausenden anderen Neuronen in ihrer Umgebung Signale. Ob diese Eingänge jedoch in der Lage sind, ein Aktionspotential auszulösen oder nicht, hängt von der Summation dieser Eingänge ab. Summation kann als ein Prozess definiert werden, durch den die erregenden und hemmenden Signale zusammen in der Lage sind, ein Aktionspotential zu erzeugen oder nicht.

Es gibt zwei Arten von Summationen, das sind die zeitliche bzw. die räumliche Summation. Räumliche Summierungen können als Signale definiert werden, die von mehreren gleichzeitigen Eingängen stammen. Die zeitliche Summierung hingegen kommt von wiederholten Eingaben. Um ein Aktionspotential zu erreichen, muss im Wesentlichen die Schwellenspannung erreicht werden. Diese kann durch Addition der einzelnen Eingänge aus zeitlicher und räumlicher Summation bestimmt werden.
Es gibt zwei Arten von Neurotransmittern, die von präsynaptischen Neuronen freigesetzt werden. Durch exzitatorische Neurotransmitter wird eine Depolarisation hervorgerufen, dies wird als exzitatorisches postsynaptisches Potential (EPSP) bezeichnet. Inhibitorische Neurotransmitter verursachen eine Hyperpolarisation, also ein inhibitorisches postsynaptisches Potenzial (IPSP). Neuronen sind in der Lage, sich gegenseitig auf verschiedene Weise zu beeinflussen. Sie sind in der Lage, Erregung und Hemmung zu bewirken und können sich gegenseitig in ihrer Erregbarkeit beeinflussen.
Defacilitation ist die Entfernung von erregenden Inputs, die einen Signalweg erleichtern können. Enthemmung hingegen ist die Entfernung hemmender Eingänge.
Mehrere Quelleneingänge an einem Neuron können räumlich summiert werden, allerdings müssen die Eingänge eng beieinander liegen, damit keiner der frühen Eingänge abfällt. Wenn ein Neuron mehrere Eingänge von einer einzigen Quelle in dem erforderlichen engen zeitlichen Abstand erhält, so dass kein Eingang abklingt, dann können diese Eingänge zeitlich summiert werden.
Ein weiterer sehr wichtiger Faktor, der bestimmt, ob das Schwellenpotential erreicht wird und ein Aktionspotential zustande kommt oder nicht, ist der Abstand zwischen der Synapse und dem neuronalen Zellkörper. Je geringer der Abstand zwischen Synapse und Zellkörper ist, desto größer ist die Summation und desto wahrscheinlicher ist es, dass ein Aktionspotenzial ausgelöst wird.
Wie wir wissen, ist das Transportmittel für ein postsynaptisches Neuron ein Dendrit. Diese Dendriten haben nur wenige spannungsgesteuerte Ionenkanäle. Daher schwächt sich das postsynaptische Potenzial beim Erreichen des neuronalen Zellkörpers ab. Dieser neuronale Zellkörper summiert diese ankommenden Potentiale. Bei der Weiterleitung des Nettopotentials zum Axonhügel wird dann ein Aktionspotential ausgelöst.
Räumliche Summation
Wenn Eingänge von mehreren Neuronen ein Aktionspotential auslösen, spricht man von räumlicher Summation. Diese Potentiale kommen meist von Dendriten, wir addieren diese Eingänge zusammen, um die räumliche Summation zu erhalten.
Je größer die Anzahl der erregenden postsynaptischen Potenziale ist, desto größer ist die Wahrscheinlichkeit, dass das Potenzial das Schwellenpotenzial erreicht, um ein Aktionspotenzial auszulösen. Umgekehrt gilt: Je größer die Anzahl der hemmenden postsynaptischen Potenziale, desto geringer ist die Chance, dass das Potenzial das Schwellenpotenzial erreicht, um ein Aktionspotenzial zu erzeugen.
Die Chancen, ein Aktionspotential auszulösen, werden auch maßgeblich davon beeinflusst, wie nah der dendritische Eingang am Axonhügel liegt. Je näher der dendritische Eingang am Axonhügel liegt, desto wahrscheinlicher ist es, dass er ein Aktionspotential auslöst. Shunting eines exzitatorischen postsynaptischen Potentials ist die Aufhebung eines exzitatorischen Inputs durch die räumliche Summation von inhibitorischen Inputs.
Temporale Summation
Wenn eine große Menge von präsynaptischen Neuronen-Aktionspotentialen postsynaptische Aktionspotentiale auslöst, die sich miteinander summieren, spricht man von temporaler Summation.
In diesem Fall ist das Intervall zwischen den Aktionspotentialen kleiner als die Dauer des postsynaptischen Aktionspotentials. Die Summation kann erhöht werden, wenn die Zeitkonstante der Zellmembran lang genug ist. Wenn das nächste postsynaptische Potenzial beginnt, summiert sich die Amplitude des vorherigen postsynaptischen Potenzials mit ihm und erzeugt ein größeres Potenzial, was die Wahrscheinlichkeit erhöht, das Schwellenpotenzial zu erreichen.
Mechanismus
Die postsynaptischen Zellen enthalten Ionenkanäle, diese Ionenkanäle können sich öffnen oder schließen, je nachdem, welcher Neurotransmitter an die Rezeptoren bindet. Das Öffnen/Schließen dieser Kanäle erzeugt ein postsynaptisches Potenzial. Es gibt 2 Arten von postsynaptischen Potenzialen. Ein exzitatorisches postsynaptisches Potenzial ist dasjenige, das die Wahrscheinlichkeit der Auslösung eines Aktionspotenzials erhöht. Ähnlich verhält es sich mit hemmenden postsynaptischen Potenzialen, die die Wahrscheinlichkeit der Auslösung eines Aktionspotenzials verringern.
Erregende Neurotransmitter (Glutamat)
Ein Paradebeispiel für einen erregenden Neurotransmitter ist Glutamat. Dieses Glutamat bindet an AMPA-Rezeptoren an der postsynaptischen Membran. Diese Bindung bewirkt einen Natrium-Kationen-Einstrom. Dieser Natriumeinstrom verursacht eine Depolarisation. Dies wird als exzitatorisches postsynaptisches Potenzial (EPSP) bezeichnet. Es ist wichtig zu beachten, dass für die EPSP-Summierung zum Erreichen des Schwellenpotentials eine große Anzahl dieser Eingänge erforderlich ist. Die Wirkung von Neurotransmittern hält viel länger an als die von präsynaptischen Impulsen.
Der Unterschied zwischen exzitatorischen postsynaptischen Potenzialen und Aktionspotenzialen besteht darin, dass exzitatorische postsynaptische Potenziale in der Lage sind, ihre Eingänge zu summieren und so eine abgestufte Reaktion zu erzeugen, im Gegensatz zur Alles-oder-Nichts-Reaktion, bei der das Schwellenpotenzial erreicht werden kann, indem ein Aktionspotenzial stimuliert wird, oder es kann überhaupt nicht erreicht werden.
Inhibitorische Neurotransmitter (GABA)
GABA ist der wichtigste Neurotransmitter, der an inhibitorischen postsynaptischen Potentialen (IPSP) beteiligt ist. Bei der Bindung an postsynaptische Neuronenrezeptoren öffnet GABA spezifische Ionenkanäle, die sich von denen unterscheiden, die beim EPSP durch exzitatorische Neurotransmitter wie Glutamat geöffnet werden. Diese Kanäle ermöglichen einen Einstrom von negativ geladenen Anionen oder einen Ausstrom von positiv geladenen Kationen. Das Anion ist in diesem Fall ein Chlorid-Ion. Die ausfließenden Kationen sind Kalium-Ionen. Beide Ionen haben den gleichen Effekt, indem sie das Membranpotential senken und damit eine Hyperpolarisation des postsynaptischen Neurons bewirken.
Die Summe dieser IPSPs und der Abfall der Membranspannung führt zu einer Abweichung vom Schwellenpotential, wodurch ein Aktionspotential gehemmt wird. Diese IPSPs und EPSPs können jedoch gleichzeitig auftreten, so dass das postsynaptische Neuron exzitatorische Signale von Glutamat und inhibitorische Signale von GABA erhalten kann. Das Ziel von hemmendem Glutamat ist es, das Membranpotential durch Hyperpolarisation vom Schwellenpotential weg zu senken.
Algebraische Verarbeitung von EPSPs und IPSPs
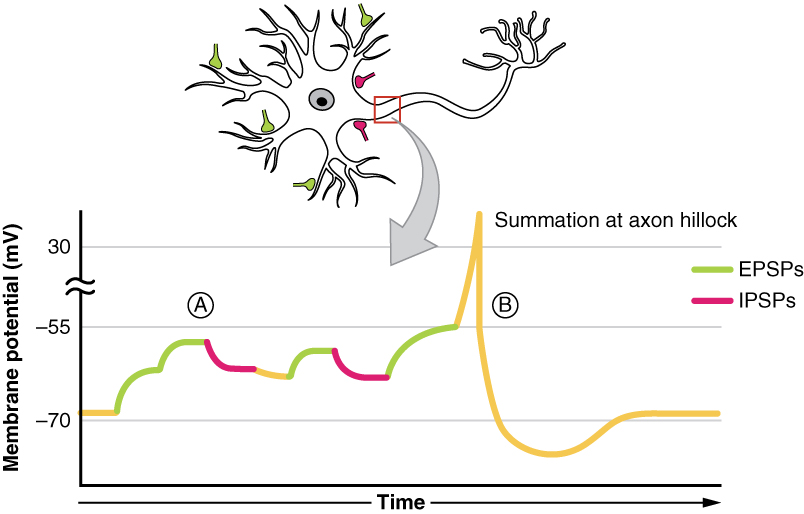
In jedem Neuron werden zu jedem Zeitpunkt zahlreiche EPSP- und IPSP-Eingänge gleichzeitig empfangen. Um den Ausgang zu bestimmen, ob das Schwellenpotential erreicht wird und ein Aktionspotential ausgelöst wird oder nicht, muss die algebraische Verarbeitung dieser EPSPs und IPSPs genommen werden. Diese Neuronen erhalten zahlreiche Eingänge, sei es von mehreren Neuronen (räumliche Summierung) oder mehrere Eingänge von einem einzelnen Neuron (zeitliche Summierung).
Dieser Ausgang hängt von der Anzahl jedes Neurotransmittertyps ab, sei es von exzitatorischen Neurotransmittern wie Glutamat, die einen Natriumioneneinstrom durch Natriumionenkanäle verursachen, oder von inhibitorischen Neurotransmittern wie GABA, die einen Chloridioneneinstrom oder Kaliumioneneinstrom durch Chlorid- bzw. Kaliumionenkanäle verursachen. Diese Synapse kann als Entscheidungspunkt bezeichnet werden, an dem die algebraische Verarbeitung dieser IPSPs und EPSPs den Ausgang bestimmt.
Axon Hillock

Der Teil des Zellkörpers eines Neurons, der mit dem Axon verbunden ist, wird Axon Hillock genannt. Er hat eine spärliche Verteilung von Nissl-Substanz. Wir können ihn durch Lichtmikroskopie identifizieren. Da dieser Axonhügel das Axon und das Soma des Neurons verbindet, ist er die letzte Region des Somas, in der die Summierung von Membranpotentialen aus synaptischen Eingängen stattfindet. Diese Summation wird dann an das Axon weitergeleitet.
Obwohl in der Vergangenheit viele glaubten, dass der Axonhügel die Auslösezone ist, in der Aktionspotentiale initiiert werden, glaubt man heute, dass das anfängliche Segment zwischen dem ersten unmyelinisierten Axonsegment und dem Axonhügelgipfel der Ort ist, an dem das Aktionspotential initiiert wird. Der positive Punkt des Axons, an dem das Aktionspotential gestartet wird, variiert von Zelle zu Zelle.
Auch hormonelle Stimulation und Neurotransmitter-Second-Messenger-Effekte können diesen positiven Punkt verändern. Die Lokalisierung von Membranpotentialen auf den somalen oder axonalen Teil der Zelle kann durch die Abgrenzung von unterschiedlichen Membrandomänen zwischen Axon und Zellkörper erreicht werden. Dies kann durch den Axonhügel erreicht werden.
Shunting
Oftmals liegen in einem Neuron die erregenden postsynaptischen und hemmenden postsynaptischen Potentiale in einem Dendriten sehr nahe beieinander. Dies wird als Shunting bezeichnet. Neben den Dendriten kann Shunting auch im Soma einer Zelle auftreten.
Die zeitliche Summation würde besagen, dass diese exzitatorischen postsynaptischen und inhibitorischen postsynaptischen Potenziale summiert werden müssen, um zu bestimmen, ob das Schwellenpotenzial erreicht und ein Aktionspotenzial ausgelöst wird oder nicht. Bei dieser Abfolge von Ereignissen im Soma eines Rufs wird jedoch der Zellwiderstand durch den inhibitorischen Input verändert. Die Zelle beginnt zu lecken, wodurch ein Shunt entsteht, anstatt die Wirkung des erregenden Inputs zu beseitigen.
Therapeutische Anwendung
Bei der Diskussion über nozizeptive Stimulation ist die zeitliche Summation die Integration sich wiederholender schmerzhafter Reize. Räumliche Summation hingegen ist die Integration von nozizeptivem Input aus großen Arealen. Viele chronische Krankheiten haben Symptome von lang anhaltenden und weit verbreiteten Schmerzen. Daher finden sich sowohl zeitliche als auch räumliche nozizeptive Summationen bei chronischen Krankheiten. Druckstimulationsexperimente liefern den Beweis dafür, dass die zeitliche Summierung von nozizeptiven Eingängen durch die räumliche Summierung erleichtert wird. Daher ist die beste Vorgehensweise bei der Behandlung chronischer Schmerzen, die Behandlung sowohl auf die räumliche als auch auf die zeitliche Summierung des Schmerzes auszurichten.
Forschung
Die meisten Experimente zur räumlichen Summierung werden an optischen und sensorischen Neuronen durchgeführt, da diese einen konstanten Frequenzbereich von erregenden und hemmenden Neuronen aufweisen. Die postsynaptische Potentialdämpfung von Neuronenzellkörpern und Dendriten ist ein großer Schwerpunkt der neuronalen Summation in den Studien der letzten Tage. Aufgrund der Tatsache, dass die Antwort dieser Interaktionen geringer ist als die Summe der Einzelantworten, werden diese Antworten als nichtlinear beschrieben. Shunting ist eine häufige Ursache für diesen Effekt. Shunting ist ein verminderter Leitwert des exzitatorischen postsynaptischen Potenzials.
Naoki Kogo und Michael Ariel konnten in ihren Experimenten am basalen Sehnervenkern der Schildkröte Informationen über die Shunting-Inhibition ableiten. Laut ihrer Arbeit verursachte die räumliche Summierung von EPSP und IPSP eine Abschwächung der exzitatorischen Antwort während der inhibitorischen Antwort. Es wurde auch festgestellt, dass nach der Abschwächung eine Verstärkung der exzitatorischen Antwort auftrat. Als Kontrolle für dieses Experiment wurde die Dämpfung getestet, wenn ein Hyperpolarisationsstrom diese spannungsempfindlichen Kanäle aktivierte. Es wurde gefolgert, dass die Dämpfung durch die Öffnung der synaptischen Rezeptorkanäle und nicht durch Hyperpolarisation verursacht wird.
Schlussfolgerung
Zu jedem Zeitpunkt empfängt ein Neuron mehrere Eingänge von mehreren verschiedenen Neuronen.
Diese Eingänge können erregend oder hemmend sein, aber um zu bestimmen, ob das Schwellenpotential erreicht wird und ein Aktionspotential ausgelöst wird oder nicht, muss eine Reihe von Faktoren betrachtet werden.
Die verschiedenen Neurotransmitter haben unterschiedliche Wirkungen. Exzitatorische Neurotransmitter wie Glutamat bewirken die Öffnung von Natriumionenkanälen, was zu einem Einstrom von Natriumionen führt und das Membranpotential erhöht. Hemmende Neurotransmitter wie GABA können Chlorid-Ionen-Kanäle öffnen, die einen Chlorid-Ionen-Einstrom verursachen, oder sie können Kalium-Ionen-Kanäle öffnen, die einen Kalium-Ionen-Efflux verursachen.
Beides senkt das Membranpotential und weicht damit von der Schwellenspannung ab, was die Wahrscheinlichkeit der Auslösung eines Aktionspotentials verringert.
Ein weiterer Faktor, der die Chancen für die Auslösung eines Aktionspotentials erhöht, ist die Nähe des dendritischen Eingangs zum Axonhügel, je näher, desto höher die Chancen für die Auslösung eines Aktionspotentials. Ein Axonhügel ist ein Teil des Somas eines Neurons, der es mit dem Axon verbindet.
Erregende Neurotransmitter erhöhen die Wahrscheinlichkeit einer Depolarisation, die Änderung der Membranspannung wird als exzitatorisches postsynaptisches Potenzial (EPSP) bezeichnet. Inhibitorische Neurotransmitter erhöhen die Wahrscheinlichkeit einer Hyperpolarisation, diese Änderung des Membranpotentials wird als inhibitorisches postsynaptisches Potential (IPSP) bezeichnet.
Ob die Schwellenspannung erreicht wird und ein Aktionspotential ausgelöst wird oder nicht, hängt von der algebraischen Summierung der einzelnen Eingänge ab. Es gibt zwei Arten der Summation.
Die Summation mehrerer Eingänge von mehreren verschiedenen präsynaptischen Neuronen, die ein Aktionspotential auslösen, wird als räumliche Summation bezeichnet. Eine hohe Frequenz von Eingängen von einem präsynaptischen Neuron, die summiert werden, um ein Aktionspotential zu erzeugen, wird als zeitliche Summation bezeichnet. Die Dauer dieses postsynaptischen Potenzials ist länger als die Dauer zwischen den jeweiligen Aktionspotenzialen.