Inleiding
De hydratatie van kooldioxide (CO2) en het mechanisme daarvan in levende systemen zijn van fundamenteel belang voor de bio-anorganische scheikunde. In 1932 werd het bestaan vastgesteld van een enzym dat CO2-hydratatie in rode bloedcellen katalyseert,31 het enzym kreeg de naam koolzuuranhydrase (afgekort CA). In 1939 werd vastgesteld dat het enzym zink (Zn) bevat.32 Omdat CO2 ofwel het beginpunt is van de fotosynthese ofwel het eindpunt van de oxidatie van substraten, is nu bekend dat koolzuuranhydrases alomtegenwoordig zijn en voorkomen in dieren, planten, bacteriën en schimmels. Verschillende enzymen uit verschillende bronnen, die dezelfde reactie katalyseren en gewoonlijk homologe structuren hebben, worden isoenzymen genoemd. Tot dusver zijn op basis van het organisme in totaal 7 verschillende klassen van CA’s geïdentificeerd: alpha, beta, gamma, delta, zeta, eta, en theta. Elke klasse kan meerdere isoenzymen bevatten. Soms heeft hetzelfde organisme meer dan één isoenzym voor een bepaalde functie, zoals het geval is bij het menselijke koolzuuranhydrase. De mens heeft 15 CA’s die tot de alfa-klasse behoren; deze isozymes verschillen naar plaats in het lichaam en naar katalytische activiteit. CA is een klassiek voorbeeld van een hydrolytisch enzym, een enzym dat de toevoeging of verwijdering van water aan een substraatmolecuul katalyseert. Meer in het bijzonder katalyseert CA de omkeerbare omzetting van kooldioxide (CO2) in bicarbonaat (HCO3-), ook wel koolzuur genoemd.

Hoewel de hydratatie van CO2 spontaan plaatsvindt in water bij pH 7, is de reactie kinetisch traag (k = 10-1 s-1), te traag om alle CO2 om te zetten die bij de ademhaling wordt geproduceerd. Pas boven pH 9 wordt de ongekatalyseerde reactie snel, dankzij de directe aanval van OH-, dat een veel beter nucleofiel is dan H2O (k = 104 M-1 s-1, waarbij M-1 verwijst naar de OH- concentratie). De figuur hieronder vergelijkt de nucleofiele aanval van water versus hydroxide (OH-) op CO2.
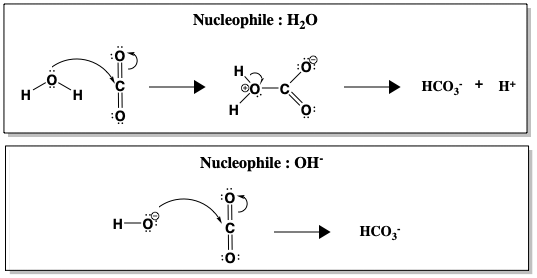
Tussen H2O en OH- verloopt de vorming van HCO3- sneller wanneer OH- het nucleofiel is. Een snellere reactie bij een hogere pH, wanneer er meer OH- ionen aanwezig zijn, suggereert dat OH- betrokken is bij de snelheidsbepalende stap. Maar realistisch gezien kan de pH van menselijk bloed niet veranderd worden om de hydratatie van CO2 te versnellen. In plaats daarvan gebruiken mensen koolzuuranhydrase om de reactie te katalyseren. Wanneer CA aanwezig is, wordt de reactie versneld tot een snelheid van k = 106 s-1.
De alomtegenwoordigheid van CA in verschillende organismen weerspiegelt het belang van deze enzymen voor de instandhouding van het leven. De snelheid van CA-gekatalyseerde CO2-hydratatie is essentieel om aan de behoeften van levende cellen te voldoen. Enkele fysiologische functies van CA zijn pH-regeling, afscheiding van elektrolyten, ionentransport en CO2-homeostase. In het spijsverteringskanaal spelen CA’s een rol bij de afscheiding van zuur en houden ze het speeksel neutraal door de pH te moduleren.G,H Van deze functies speelt CA met name een rol bij het transport van CO2 en HCO3- gerelateerd aan de ademhaling, het proces van atmosferische zuurstof- en kooldioxide-uitwisseling dat plaatsvindt wanneer mensen zuurstof inademen en kooldioxide uitademen. Omdat CO2 slecht oplosbaar is, moet het worden omgezet in een beter oplosbare vorm, HCO3-, om door het lichaam te worden getransporteerd. Bicarbonaat-ion (HCO3-) bereikt uiteindelijk de longen, wordt weer omgezet in CO2, en verlaat het lichaam door uitademing.C
Medisch onderzoek rond CA richt zich op de Zn-bevattende actieve site als therapeutisch doelwit voor verschillende ziektebehandelingen; zowel CA-remmers als -activators worden in het geneesmiddelenontwerp opgenomen. CA-remmers worden gebruikt als behandeling voor epilepsie, maagzweren, kanker, zwaarlijvigheid en andere neurologische aandoeningen. In het oog produceert CA waterstofionen die de optische druk in stand houden. Te veel druk in het oog kan echter de oogzenuw beschadigen en glaucoom veroorzaken. CA-activiteit kan een concentratiegradiënt creëren die het transport van water naar de oogzenuw stimuleert. Wanneer zich te veel water rond de oogzenuw bevindt, neemt de druk rond de zenuw toe en ontstaat schade. Remming van CA is een belangrijke behandeling van glaucoom geworden.
Naast farmaceutische toepassingen is CA ook onderzocht op zijn nut voor koolstofvastlegging en koolstofsensortoepassingen. Koolstofvastlegging en -opslag vindt plaats wanneer CA CO2 omzet in bicarbonaat. Een verhoogde beschikbaarheid van bicarbonaat in de aanwezigheid van calciumionen (Ca2+) veroorzaakt de neerslag van calciet (CaCO3). Dit proces, dat bio-mineralisatie wordt genoemd, kan een levensvatbare methode zijn om CO2 op lange termijn op te slaan in calciet en zo de CO2-uitstoot te beperken.
De details over de structuur en functie van de Zn-bevattende actieve site van CA zijn in 80 jaar onderzoek opgehelderd. Het huidige artikel gaat dieper in op de actieve site van het metalloenzym met behulp van bioinorganische concepten.
Old Text Below ————————————————————————————————————————————————————————————————–
Koolstofdioxidehydratie en het mechanisme daarvan in levende systemen zijn van fundamenteel belang voor de bioinorganische chemie. In 1932 werd het bestaan vastgesteld van een enzym dat CO2-hydratatie in rode bloedcellen katalyseert,31 Het enzym kreeg de naam koolzuuranhydrase (afgekort CA). In 1939 werd vastgesteld dat het enzym zink bevat.32 Omdat CO2 ofwel het beginpunt is van de fotosynthese ofwel het eindpunt van de oxidatie van substraten, is nu bekend dat koolzuuranhydrases alomtegenwoordig zijn en voorkomen in dieren, planten en verscheidene bacteriën. Verschillende enzymen uit verschillende bronnen, die dezelfde reactie katalyseren en gewoonlijk homologe structuren hebben, worden isoenzymen genoemd. Soms heeft hetzelfde organisme meer dan een isoenzym voor een bepaalde functie, zoals het geval is bij het menselijke koolzuuranhydrase.
CO2-gas is relatief goed oplosbaar in water (3 x 10-2 M bij kamertemperatuur onder pCO2 = 1 atm), waarbij het in evenwicht komt met waterstofcarbonaat bij pKa 6.1:
De ongekatalyseerde reactie is kinetisch traag rond fysiologische pH (k \(\simeq\) 10-1 s-1), terwijl in aanwezigheid van het meest efficiënte isoenzym van CA, het maximale CO2-omzetgetal (d.w.z, het aantal substraatmoleculen dat per tijdseenheid door elk enzymmolecuul wordt omgezet)33 106 s-1 bedraagt. De ongekatalyseerde aanval door water op CO2 kan worden vergemakkelijkt door twee waterstofgebonden watermoleculen, waarvan de ene de koolstof activeert door middel van een waterstofbrug aan een eindzuurstof van CO2, en de andere het koolstofatoom bindt via zuurstof: 34,35
(\tag{2.7})
Pas boven pH 9 wordt de ongekatalyseerde reactie snel, door de directe aanval van OH-, dat een veel beter nucleofiel is dan H2O (k \(\simeq\) 104 M-1s-1, waarbij M-1 verwijst naar de OH- concentratie):
Anderzijds is de snelheidsconstante in aanwezigheid van het enzym, kcat genaamd, pH-onafhankelijk boven pH 8 bij elk CA-isoenzym (figuur 2.2).33,36
In vitro is koolzuuranhydrase zeer veelzijdig en katalyseert het verschillende reacties waarbij zowel OH- als H+ betrokken zijn, zoals de hydrolyse van esters en de hydratatie van aldehyden. De verschillende isoenzymen zijn in verschillende mate van verfijning gekarakteriseerd. De hoog-actieve vormen worden aangeduid met II (kcat \(\simeq\) 106 s-1 bij 25 °C); de laag-actieve vormen I (kcat \(\simeq\) 105 s-1), en de zeer-laag-actieve vormen III (kcat \(\simeq\) 103 s-1).37 Er is röntgenstructuurinformatie met een nominale resolutie van 2 Å beschikbaar voor HCA I38 en HCA II,39 waarbij H staat voor mens. De structuur van HCA II is onlangs verfijnd.40 Er worden hoge-resolutie structuren gerapporteerd van mutanten en van hun substraat- en inhibitorderivaten.211 Alle isoenzymen zijn eenkernige polypeptiden, met een M.W. van ongeveer 30 kDa en één zinkion per molecuul. Ze hebben de vorm van een rugbybal met een spleet van 16 Å diep die door de zuidpool loopt (figuur 2.3 Zie kleurenplaatgedeelte, blz. C-2.). Op de bodem van de spleet is het zinkion verankerd aan het eiwit door drie histidine stikstofatomen en blootgesteld aan oplosmiddel. Twee histidines (His-94 en His-96, HCA I-nummering) zijn aan zink gebonden via hun N(\(\)2 atomen, terwijl één (His-119) is gebonden via zijn N\(\delta)1 atoom (figuur 2.4). Het is vrij algemeen dat histidines zink even goed binden via elk van de twee histidine-nitrogenen, waarbij de voorkeur waarschijnlijk wordt bepaald door de sterische beperkingen die door de eiwitvouwing worden opgelegd. De drie NH-protonen van histidine zijn alle betrokken bij H-bindingen (figuur 2.4). Histidine-119 is betrokken bij H-bonding met een glutamaat residu. Zoals gezegd, zou dit een manier kunnen zijn om de basiciteit van de metaalliganden te controleren. Een oplosmiddelmolecuul gebonden aan zink is betrokken bij een H-binding met Thr-199, dat op zijn beurt weer een H-binding heeft met Glu-106. Dit H-bonding netwerk is belangrijk voor het begrijpen van de subtiele structuurveranderingen die optreden bij pH veranderingen; deze zouden in principe de pH-afhankelijke eigenschappen kunnen verklaren. Hoewel de structuur van kristallen gegroeid bij pH 8 in sulfaat-bevattende buffer enige aanwijzing geeft van een enkele oplosmiddelmolecule gebonden aan zink (Figuren 2.3 en 2.5 Zie kleurenplaat sectie, pagina C2, C3.), geven theoretische studies aan dat twee watermoleculen op bindingsafstand kunnen zijn.42 Een dergelijke bevinding is consistent met spectroscopische studies aan andere derivaten en met het concept dat binding en onthechting van substraten plaatsvindt door middel van vijf coördinatie.
Net als bij elk zinkenzym waarin zink zich op de katalytische plaats bevindt, gaat de activiteit verloren als het metaal wordt verwijderd, en wordt deze hersteld door zinkopname. De tertiaire structuur van koolzuuranhydrase blijft behouden bij afwezigheid van zink; zelfs het gedenatureerde apoproteïne kan zich spontaan omvouwen van een willekeurige spoel tot een natieve conformatie. Hoewel een dergelijk proces versneld wordt door zink,43,44 lijkt de aanwezigheid van het metaal geen absolute vereiste te zijn voor de correcte vouwing van CA, terwijl het een absolute vereiste is voor verscheidene andere metalloproteïnen.23 ,29,30
Anionen worden in de metaalholte aangetrokken door het positieve Zn(N3OH2)2+ gedeelte, en men denkt dat ze zich in koolzuuranhydrase zeer effectief aan zink binden; het gebruik ervan moet dus zoveel mogelijk worden vermeden als het doel is het enzym te bestuderen zoals het is. Wanneer het eiwit wordt gedialyseerd tegen vers dubbel gedestilleerd of zorgvuldig gedeïoniseerd water onder een inerte atmosfeer, nadert de pH van het monster het isoëlektrische punt, dat voor HCA I en boviene (BCA II) enzymen onder 6 ligt. De pH kan vervolgens worden aangepast door passende toevoegingen van NaOH. Alle in de literatuur vermelde metingen die in acetaat-, fosfaat-, imidazool- of trisulfaatbuffers zijn uitgevoerd, worden beïnvloed door de interferentie van het anion met het metaalion. Buffersoorten die grote anionen bevatten, zoals Hepes (4ethaansulfonzuur), kunnen echter wel worden gebruikt,45 aangezien deze anionen de holte niet binnendringen.
Er zijn veel aanwijzingen dat zink in de hoge-pH-vorm van CA vier-gecoördineerd is met een OH-groep in de vierde coördinatieplaats. Bij lage pH bestaat het enzym in een vorm die gecoördineerd water bevat; het coördinatiegetal kan vier (één watermolecule) of vijf (twee watermoleculen) zijn. Uiteraard hangt het voorkomen van de lage-pH-soorten af van de pKa’s van de complexe zuur-base-evenwichten.