Introduzione
L’idratazione del biossido di carbonio (CO2) e il suo meccanismo nei sistemi viventi sono di fondamentale importanza per la chimica bioinorganica. Nel 1932 fu stabilita l’esistenza di un enzima che catalizzava l’idratazione della CO2 nei globuli rossi,31 L’enzima fu chiamato anidrasi carbonica (abbreviato CA). Nel 1939 si riconobbe che l’enzima conteneva zinco (Zn).32 Poiché la CO2 è il punto di partenza per la fotosintesi o il punto finale dell’ossidazione del substrato, le anidrasi carboniche sono ora note per essere ubiquitarie, presenti in animali, piante, batteri e funghi. Enzimi diversi da fonti diverse, che catalizzano la stessa reazione e di solito hanno strutture omologhe, sono chiamati isoenzimi. Finora, un totale di 7 classi distinte di CA sono state identificate in base all’organismo: alfa, beta, gamma, delta, zeta, eta e theta. Ogni classe può contenere più isoenzimi. A volte lo stesso organismo ha più di un isoenzima per una particolare funzione, come è vero per l’anidrasi carbonica umana. Gli esseri umani hanno 15 CA che appartengono alla classe alfa; questi isozimi variano in base alla posizione nel corpo e all’attività catalitica. CA è un classico esempio di enzima idrolitico, che catalizza l’aggiunta o la rimozione di acqua da una molecola di substrato. Più specificamente, CA catalizza la conversione reversibile dell’anidride carbonica (CO2) in bicarbonato (HCO3-), detto anche acido carbonico.

Anche se l’idratazione della CO2 è spontanea in acqua a pH 7, la reazione è cineticamente lenta (k = 10-1 s-1), troppo lenta per convertire tutta la CO2 prodotta nella respirazione. Solo sopra il pH 9 la reazione non catalizzata diventa veloce, a causa dell’attacco diretto di OH-, che è un nucleofilo molto migliore di H2O (k = 104 M-1 s-1, dove M-1 si riferisce alla concentrazione di OH-). La figura qui sotto confronta l’attacco nucleofilo dell’acqua rispetto all’idrossido (OH-) sulla CO2.
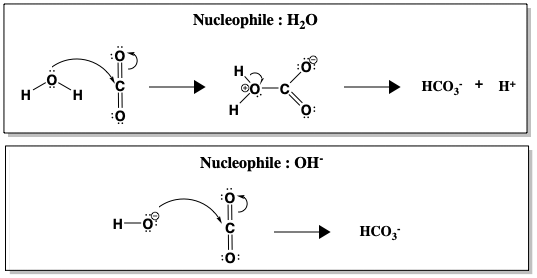
Tra H2O e OH-, la formazione di HCO3- avviene più velocemente quando OH- è il nucleofilo. Una reazione più veloce a pH più alto, quando sono presenti più ioni OH-, suggerisce che OH- è coinvolto nella fase di determinazione del tasso. Tuttavia, realisticamente, il pH del sangue umano non può essere cambiato per accelerare l’idratazione della CO2. Invece, gli esseri umani usano l’anidrasi carbonica per catalizzare la reazione. Quando CA è presente, la reazione è accelerata ad un tasso di k = 106 s-1.
L’ubiquità di CA in diversi organismi riflette l’importanza di questi enzimi nel sostenere la vita. La velocità di idratazione della CO2 catalizzata da CA è essenziale per soddisfare le esigenze delle cellule viventi. Alcune funzioni fisiologiche di CA includono la regolazione del pH, la secrezione di elettroliti, il trasporto di ioni e l’omeostasi di CO2. Nel tratto digestivo, i CA svolgono un ruolo nella secrezione di acido e mantengono la saliva neutra modulando il pH.G,H Tra queste funzioni, i CA svolgono soprattutto un ruolo nel trasporto di CO2 e HCO3- legato alla respirazione, il processo di scambio di ossigeno atmosferico e anidride carbonica che avviene quando gli esseri umani inspirano ossigeno ed espirano anidride carbonica. Con una bassa solubilità, la CO2 deve essere convertita in una forma più solubile, HCO3-, per il trasporto in tutto il corpo. Lo ione bicarbonato (HCO3-) alla fine raggiunge i polmoni, viene riconvertito in CO2 ed esce dal corpo attraverso l’espirazione.C
La ricerca medica che ruota intorno a CA si concentra sul sito attivo contenente Zn come bersaglio terapeutico per vari trattamenti di malattie; sia gli inibitori che gli attivatori di CA sono incorporati nella progettazione di farmaci. Gli inibitori di CA sono usati come trattamento per l’epilessia, le ulcere, il cancro, l’obesità e altri disturbi neurologici. Nell’occhio, il CA produce ioni idrogeno che mantengono la pressione ottica. Tuttavia, troppa pressione nell’occhio può danneggiare il nervo ottico e causare il glaucoma. L’attività del CA può creare un gradiente di concentrazione che guida il trasporto di acqua verso il nervo ottico. Quando troppa acqua si trova intorno al nervo ottico, la pressione intorno al nervo aumenta causando danni. L’inibizione di CA è diventata un trattamento chiave del glaucoma.
Oltre alle applicazioni farmaceutiche, il CA è stato anche studiato per la sua utilità nella cattura del carbonio e nelle applicazioni dei sensori di carbonio. La cattura e lo stoccaggio del carbonio avviene quando i CA convertono la CO2 in bicarbonato. La maggiore disponibilità di bicarbonato in presenza di ioni di calcio (Ca2+) causa la precipitazione di calcite (CaCO3). Questo processo, chiamato bio-mineralizzazione, può essere una modalità praticabile di stoccaggio a lungo termine di CO2 nella calcite per mitigare le emissioni di CO2.
I dettagli sulla struttura e la funzione del sito attivo di CA contenente Zn sono stati chiariti in 80 anni di ricerca. L’articolo attuale approfondisce il sito attivo del metalloenzima usando concetti bioinorganici.
Testo vecchio sotto ————————————————————————————————————————————————————————————————–
L’idratazione del biossido di carbonio e il suo meccanismo nei sistemi viventi sono di fondamentale importanza per la chimica bioinorganica. Nel 1932 fu stabilita l’esistenza di un enzima che catalizzava l’idratazione della CO2 nei globuli rossi,31 L’enzima fu chiamato anidrasi carbonica (abbreviato CA). Nel 1939 si riconobbe che l’enzima conteneva zinco.32 Poiché la CO2 è il punto di partenza per la fotosintesi o il punto finale dell’ossidazione del substrato, le anidrasi carboniche sono ora note per essere ubiquitarie e si trovano negli animali, nelle piante e in diversi batteri. Enzimi diversi da fonti diverse, che catalizzano la stessa reazione e di solito hanno strutture omologhe, sono chiamati isoenzimi. A volte lo stesso organismo ha più di un isoenzima per una particolare funzione, come è vero per l’anidrasi carbonica umana.
Il gas CO2 è relativamente solubile in acqua (3 x 10-2 M a temperatura ambiente sotto pCO2 = 1 atm), in equilibrio con l’idrogeno carbonato a pKa 6.1:
La reazione non catalizzata è cineticamente lenta intorno al pH fisiologico (k \(\simeq\) 10-1 s-1), mentre, in presenza dell’isoenzima più efficiente della CA, il numero massimo di turnover della CO2 (cioè, il numero di molecole di substrato trasformate per unità di tempo da ogni molecola di enzima)33 è \(\simeq\) 106 s-1. L’attacco non catalizzato dell’acqua sulla CO2 può essere facilitato da due molecole d’acqua con legame a idrogeno, una delle quali attiva il carbonio per mezzo di un legame a idrogeno con un ossigeno terminale della CO2, l’altra delle quali lega l’atomo di carbonio tramite l’ossigeno: 34,35
(\tag{2.7})
Solo sopra il pH 9 la reazione non catalizzata diventa veloce, a causa dell’attacco diretto di OH-, che è un nucleofilo molto migliore di H2O (k \(\simeq\) 104 M-1s-1, dove M-1 si riferisce alla concentrazione di OH-):
D’altra parte, la costante di tasso in presenza dell’enzima, chiamata kcat, è indipendente dal pH sopra il pH 8 in ogni isoenzima CA (Figura 2.2).33,36
In vitro, l’anidrasi carbonica è abbastanza versatile, catalizzando diverse reazioni che coinvolgono sia OH- che H+, come l’idrolisi degli esteri e l’idratazione delle aldeidi. I vari isoenzimi sono stati caratterizzati a diversi gradi di sofisticazione. Le forme ad alta attività sono etichettate come II (kcat \(\simeq\) 106 s-1 a 25 °C); le forme a bassa attività I (kcat \(\simeq\) 105 s-1), e le forme a bassissima attività III (kcat \(\simeq\) 103 s-1).37 Sono disponibili informazioni strutturali ai raggi X con risoluzione nominale di 2 Å per HCA I38 e HCA II,39 dove H indica l’uomo. La struttura di HCA II è stata raffinata di recente.40 Strutture ad alta risoluzione di mutanti e dei loro derivati di substrato e inibitori sono state riportate.211 Tutti gli isoenzimi sono polipeptidi a catena singola, con M.W. di circa 30 kDa e uno ione zinco per molecola. Hanno la forma di una palla da rugby con una fessura profonda 16 Å che attraversa il polo sud (Figura 2.3 Vedi la sezione della piastra a colori, pagina C-2.). Sul fondo della fessura, lo ione zinco è ancorato alla proteina da tre atomi di azoto di istidina ed è esposto al solvente. Due istidine (His-94 e His-96, numerazione HCA I) sono legate allo zinco attraverso i loro atomi N(\epsilon\2), mentre una (His-119) è legata attraverso il suo atomo N(\delta\1) (Figura 2.4). È abbastanza generale che le istidine leghino lo zinco ugualmente bene attraverso uno dei due nitrogeni dell’istidina, la preferenza è probabilmente dettata dai vincoli sterici imposti dal ripiegamento della proteina. I tre protoni NH dell’istidina sono tutti impegnati nel legame H (Figura 2.4). L’istidina-119 è coinvolta nel legame H con un residuo di glutammato. Come accennato, questo potrebbe essere un modo per controllare la basicità dei ligandi metallici. Una molecola di solvente legata allo zinco è coinvolta in un legame H con Thr-199, che a sua volta è legato H a Glu-106. Questa rete di H-bonding è importante per comprendere i sottili cambiamenti strutturali che si verificano con i cambiamenti di pH; questi potrebbero, in linea di principio, spiegare le proprietà dipendenti dal pH. Sebbene la struttura dei cristalli cresciuti a pH 8 in un tampone contenente solfato dia qualche indicazione di una singola molecola di solvente legata allo zinco (Figure 2.3 e 2.5 Vedi la sezione delle tavole a colori, pagine C2, C3.), studi teorici indicano che due molecole d’acqua possono essere a distanze di legame.42 Tale risultato è coerente con studi spettroscopici su altri derivati e con il concetto che l’attacco e il distacco dei substrati avvengono attraverso cinque coordinazioni.
Proprio come è vero per ogni enzima in cui lo zinco si trova nel sito catalitico, l’attività viene persa se il metallo viene rimosso, e viene ripristinata dalla ripresa dello zinco. La struttura terziaria dell’anidrasi carbonica è mantenuta in assenza di zinco; anche l’apoproteina denaturata può ripiegarsi spontaneamente da una bobina casuale a una conformazione nativa. Sebbene tale processo sia accelerato dallo zinco,43,44 la presenza del metallo non sembra essere un requisito assoluto per il corretto ripiegamento di CA, mentre è un requisito assoluto per diverse altre metalloproteine.23 ,29,30
Gli anioni sono attratti nella cavità metallica dalla parte positiva Zn(N3OH2)2+, e si ritiene che si leghino allo zinco nell’anidrasi carbonica molto efficacemente; quindi il loro uso dovrebbe essere evitato il più possibile se l’obiettivo è quello di studiare l’enzima così com’è. Quando la proteina viene dializzata contro acqua fresca bidistillata o accuratamente deionizzata in atmosfera inerte, il pH del campione si avvicina al punto isoelettrico, che è inferiore a 6 per gli enzimi HCA I e bovino (BCA II). Il pH può quindi essere regolato con opportune aggiunte di NaOH. Tutte le misurazioni riportate in letteratura eseguite in tamponi di acetato, fosfato, imidazolo o tris solfato sono influenzate dall’interferenza dell’anione con lo ione metallico. Tuttavia, le specie tampone che contengono grandi anioni come Hepes (acido 4etansolfonico) possono essere utilizzati,45 poiché questi anioni non entrano nella cavità.
Ci sono molte indicazioni che lo zinco nella forma ad alto pH di CA è quattro-coordinato con un gruppo OH nel quarto sito di coordinamento. A basso pH l’enzima esiste in una forma che contiene acqua coordinata; il numero di coordinazione può essere quattro (una molecola d’acqua) o cinque (due molecole d’acqua). Naturalmente, la presenza delle specie a basso pH dipende dai pKa degli equilibri acido-base complessi.