Wprowadzenie
Uwodnienie dwutlenku węgla (CO2) i jego mechanizm w systemach żywych mają fundamentalne znaczenie dla chemii bioorganicznej. W 1932 r. stwierdzono istnienie enzymu katalizującego hydratację CO2 w krwinkach czerwonych,31 Enzym ten nazwano anhydrazą węglową (w skrócie CA). W 1939 r. stwierdzono, że enzym ten zawiera cynk (Zn).32 Ponieważ CO2 jest albo punktem wyjścia fotosyntezy, albo punktem końcowym utleniania substratów, anhydrazy węglowe są obecnie znane jako wszechobecne, występujące u zwierząt, roślin, bakterii i grzybów. Różne enzymy pochodzące z różnych źródeł, katalizujące tę samą reakcję i zazwyczaj posiadające homologiczną strukturę, określane są mianem izoenzymów. Do tej pory zidentyfikowano łącznie 7 różnych klas CA w zależności od organizmu: alfa, beta, gamma, delta, zeta, eta i theta. Każda klasa może zawierać wiele izoenzymów. Czasami ten sam organizm posiada więcej niż jeden izoenzym dla danej funkcji, jak to ma miejsce w przypadku ludzkiej anhydrazy węglowej. Ludzie mają 15 CA należących do klasy alfa; izozymy te różnią się między sobą lokalizacją w organizmie i aktywnością katalityczną. CA jest klasycznym przykładem enzymu hydrolitycznego, takiego, który katalizuje dodawanie lub usuwanie wody do cząsteczki substratu. Dokładniej, CA katalizuje odwracalną konwersję dwutlenku węgla (CO2) do wodorowęglanu (HCO3-), zwanego również kwasem węglowym.

Ale chociaż hydratacja CO2 jest spontaniczna w wodzie przy pH 7, reakcja jest kinetycznie powolna (k = 10-1 s-1), zbyt wolna, aby przekształcić cały CO2 produkowany w procesie oddychania. Dopiero powyżej pH 9 niekatalizowana reakcja staje się szybka, dzięki bezpośredniemu atakowi OH-, który jest znacznie lepszym nukleofilem niż H2O (k = 104 M-1 s-1, gdzie M-1 odnosi się do stężenia OH-). Poniższy rysunek porównuje atak nukleofilowy wody i wodorotlenku (OH-) na CO2.
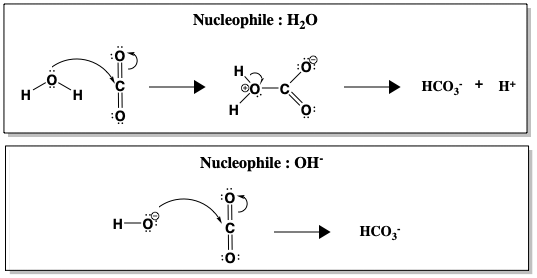
Pomiędzy H2O i OH-, tworzenie HCO3- zachodzi szybciej, gdy OH- jest nukleofilem. Szybsza reakcja przy wyższym pH, gdy obecnych jest więcej jonów OH-, sugeruje, że OH- jest zaangażowany w etap determinujący tempo. Jednakże, realistycznie rzecz biorąc, pH ludzkiej krwi nie może być zmienione, aby przyspieszyć hydratację CO2. Zamiast tego, ludzie używają anhydrazy węglowej do katalizowania reakcji. Kiedy CA jest obecny, reakcja jest przyspieszona do tempa k = 106 s-1.
Wszechobecność CA w różnych organizmach odzwierciedla znaczenie tych enzymów w podtrzymywaniu życia. Szybkość katalizowanej przez CA hydratacji CO2 jest niezbędna do zaspokojenia potrzeb żywych komórek. Niektóre fizjologiczne funkcje CA obejmują regulację pH, wydzielanie elektrolitów, transport jonów i homeostazę CO2. W przewodzie pokarmowym CAs odgrywa rolę w wydzielaniu kwasu i utrzymuje ślinę w stanie neutralnym poprzez modulację pH.G,H Wśród tych funkcji CA odgrywa przede wszystkim rolę w transporcie CO2 i HCO3- związanych z oddychaniem, procesem wymiany tlenu atmosferycznego i dwutlenku węgla, który występuje, gdy ludzie wdychają tlen i wydychają dwutlenek węgla. Ze względu na niską rozpuszczalność, CO2 musi być przekształcony w bardziej rozpuszczalną formę, HCO3-, w celu transportu w całym organizmie. Jon wodorowęglanowy (HCO3-) ostatecznie dociera do płuc, zostaje przekształcony z powrotem w CO2 i opuszcza organizm poprzez wydech.C
Badania medyczne obracające się wokół CA koncentrują się na zawierającym Zn miejscu aktywnym jako celu terapeutycznym dla różnych metod leczenia chorób; zarówno inhibitory CA, jak i aktywatory są włączone do projektowania leków. Inhibitory CA są stosowane w leczeniu epilepsji, wrzodów, raka, otyłości i innych zaburzeń neurologicznych. W oku CA wytwarza jony wodorowe, które utrzymują ciśnienie w oku. Jednak zbyt wysokie ciśnienie w oku może uszkodzić nerw wzrokowy i spowodować jaskrę. Aktywność CA może tworzyć gradient stężenia, który napędza transport wody do nerwu wzrokowego. Gdy zbyt dużo wody znajduje się wokół nerwu wzrokowego, ciśnienie wokół nerwu wzrasta, powodując jego uszkodzenie. Zahamowanie aktywności CA stało się kluczowym sposobem leczenia jaskry.
Poza zastosowaniami farmaceutycznymi, CA został również zbadany pod kątem jego użyteczności w wychwytywaniu węgla i zastosowaniach czujników węgla. Wychwytywanie i magazynowanie węgla odbywa się poprzez konwersję przez CA CO2 do wodorowęglanu. Zwiększona dostępność wodorowęglanów w obecności jonów wapnia (Ca2+) powoduje wytrącanie się kalcytu (CaCO3). Proces ten, nazywany bio-mineralizacją, może być realnym sposobem długoterminowego magazynowania CO2 w kalcycie w celu złagodzenia emisji CO2.
Szczegóły dotyczące struktury i funkcji zawierającego Zn miejsca aktywnego CA zostały wyjaśnione w ciągu 80 lat badań. W niniejszym artykule zagłębiamy się w miejsce aktywne metaloenzymu, wykorzystując koncepcje bioorganiczne.
Old Text Below ————————————————————————————————————————————————————————————————–
Hydratacja dwutlenku węgla i jej mechanizm w układach żywych mają fundamentalne znaczenie dla chemii bioorganicznej. W 1932 roku ustalono istnienie enzymu katalizującego hydratację CO2 w krwinkach czerwonych,31 Enzym ten nazwano anhydrazą węglową (w skrócie CA). W 1939 roku stwierdzono, że enzym ten zawiera cynk.32 Ponieważ CO2 jest albo punktem wyjścia fotosyntezy, albo punktem końcowym utleniania substratów, anhydrazy węglowe są obecnie znane jako wszechobecne, występujące u zwierząt, roślin i wielu bakterii. Różne enzymy pochodzące z różnych źródeł, katalizujące tę samą reakcję i zazwyczaj posiadające homologiczną strukturę, są określane mianem izoenzymów. Czasami ten sam organizm posiada więcej niż jeden izoenzym dla danej funkcji, jak to ma miejsce w przypadku ludzkiej anhydrazy węglowej.
GazowyCO2 jest względnie rozpuszczalny w wodzie (3 x 10-2 M w temperaturze pokojowej pod ciśnieniem pCO2 = 1 atm), wyrównując się z węglanem wodoru przy pKa 6.1:
Reakcja niekatalizowana jest kinetycznie powolna w okolicach fizjologicznego pH (k 10-1 s-1), natomiast w obecności najwydajniejszego izoenzymu CA, maksymalna liczba obrotu CO2 (tj, liczba cząsteczek substratu przekształcanych w jednostce czasu przez każdą cząsteczkę enzymu)33 wynosi ∙(∙simeq\) 106 s-1. Niekatalizowany atak wody na CO2 może być ułatwiony przez dwie cząsteczki wody z wiązaniami wodorowymi, z których jedna aktywuje węgiel za pomocą wiązania wodorowego z terminalnym tlenem CO2, a druga wiąże atom węgla za pośrednictwem tlenu: 34,35
\tag{2.7}})
Dopiero powyżej pH 9 niekatalizowana reakcja staje się szybka, dzięki bezpośredniemu atakowi OH-, który jest znacznie lepszym nukleofilem niż H2O (k ≥ 104 M-1s-1, gdzie M-1 odnosi się do stężenia OH-):
Z drugiej strony, stała szybkości w obecności enzymu, zwana kcat, jest niezależna od pH powyżej pH 8 w każdym izoenzymie CA (Rysunek 2.2).33,36
In vitro, anhydraza węglowa jest dość wszechstronna, katalizuje kilka reakcji, które angażują zarówno OH- jak i H+, takie jak hydroliza estrów i hydratacja aldehydów. Różne izoenzymy zostały scharakteryzowane w różnym stopniu zaawansowania. Formy o wysokiej aktywności oznaczane są jako II (kcat ≥ 106 s-1 w 25°C); formy o niskiej aktywności jako I (kcat ≥ 105 s-1), a formy o bardzo niskiej aktywności jako III (kcat ≥ 103 s-1).37 Rentgenowskie informacje strukturalne o nominalnej rozdzielczości 2 Å dostępne są dla HCA I38 i HCA II,39 gdzie H oznacza człowieka. Ostatnio udoskonalono strukturę HCA II.40 Obecnie opisywane są struktury o wysokiej rozdzielczości mutantów oraz ich substratów i pochodnych inhibitorów.211 Wszystkie izoenzymy są jednołańcuchowymi polipeptydami, o M.W. około 30 kDa i jednym jonie cynku na cząsteczkę. Mają one kształt piłki do rugby ze szczeliną o głębokości 16 Å biegnącą przez biegun południowy (Rysunek 2.3 Patrz sekcja kolorowych płytek, strona C-2.). Na dnie szczeliny jon cynku jest zakotwiczony do białka przez trzy atomy azotu histydyny i jest wystawiony na działanie rozpuszczalnika. Dwie histydyny (His-94 i His-96, numeracja HCA I) są związane z cynkiem poprzez swoje atomy N (∗epsilon) 2, podczas gdy jedna (His-119) jest związana poprzez swój atom N (∗delta) 1 (Rysunek 2.4). Jest dość powszechne, że histydyny wiążą cynk równie dobrze przez jeden z dwóch nitrogenów histydyny, przy czym preferencje są prawdopodobnie podyktowane ograniczeniami sterycznymi narzuconymi przez fałdowanie białka. Wszystkie trzy protony NH histydyny zaangażowane są w wiązania H (rysunek 2.4). Histydyna-119 jest zaangażowana w wiązanie H z resztą glutaminianową. Jak wspomniano, może to być sposób na kontrolowanie zasadowości ligandów metalicznych. Cz±steczka rozpuszczalnika zwi±zana z cynkiem jest zaangażowana w wi±zanie H z Thr-199, który z kolei jest zwi±zany H z Glu-106. Ta sieć wiązań H jest ważna dla zrozumienia subtelnych zmian strukturalnych, które zachodzą przy zmianach pH; mogłyby one, w zasadzie, odpowiadać za właściwości zależne od pH. Chociaż struktura kryształów hodowanych przy pH 8 w buforze zawierającym siarczan daje pewne wskazania na pojedynczą cząsteczkę rozpuszczalnika związaną z cynkiem (Rysunki 2.3 i 2.5 Patrz sekcja kolorowych płytek, strony C2, C3.), badania teoretyczne wskazują, że dwie cząsteczki wody mogą znajdować się w odległości wiązania.42 Takie odkrycie jest zgodne z badaniami spektroskopowymi na innych pochodnych oraz z koncepcją, że przyłączanie i odłączanie substratów zachodzi poprzez pięć koordynacji.
Tak jak w przypadku każdego enzymu cynkowego, w którym cynk znajduje się w miejscu katalitycznym, aktywność jest tracona w przypadku usunięcia metalu i przywracana przez wychwyt cynku. Struktura trzeciorzędowa anhydrazy węglowej jest zachowana pod nieobecność cynku; nawet zdenaturowana apoproteina może spontanicznie zmienić kształt z przypadkowej spirali na konformację zbliżoną do natywnej. Chociaż proces ten jest przyspieszany przez cynk,43,44 obecność tego metalu nie wydaje się być bezwzględnym wymogiem dla prawidłowego składania CA, podczas gdy jest to bezwzględny wymóg dla kilku innych metaloprotein.23 ,29,30
Aniony są przyciągane w zagłębieniu metalicznym przez dodatnią cząsteczkę Zn(N3OH2)2+ i uważa się, że wiążą się z cynkiem w anhydrazie węglowej bardzo skutecznie; więc ich stosowania należy unikać w jak największym stopniu, jeśli celem jest badanie enzymu w jego postaci. Kiedy białko jest dializowane ze świeżo podwójnie destylowaną lub starannie dejonizowaną wodą w atmosferze obojętnej, pH próbki zbliża się do punktu izoelektrycznego, który jest poniżej 6 dla enzymów HCA I i bydlęcych (BCA II). Następnie pH można skorygować przez odpowiednie dodanie NaOH. Wszystkie opisywane w literaturze pomiary wykonywane w buforach octanowych, fosforanowych, imidazolowych lub trisiarczanowych są zakłócane przez interferencję anionu z jonem metalu. Można jednak stosować gatunki buforów zawierające duże aniony, takie jak Hepes (kwas 4etanosulfonowy),45 ponieważ aniony te nie wchodzą do wnęki.
Wiele wskazuje na to, że cynk w formie CA o wysokim pH jest czterokordycyjny z grupą OH w czwartym miejscu koordynacyjnym. Przy niskim pH enzym występuje w formie zawierającej skoordynowaną wodę; liczba koordynacyjna może być czterokoronowa (jedna cząsteczka wody) lub pięciokoronowa (dwie cząsteczki wody). Oczywiście występowanie gatunków o niskim pH zależy od pKa złożonych równowag kwasowo-zasadowych.