Introduction
L’hydratation du dioxyde de carbone (CO2) et son mécanisme dans les systèmes vivants sont d’une importance fondamentale pour la chimie bioinorganique. En 1932, l’existence d’une enzyme catalysant l’hydratation du CO2 dans les globules rouges a été établie,31 L’enzyme a été nommée anhydrase carbonique (en abrégé CA). En 1939, il a été reconnu que l’enzyme contenait du zinc (Zn).32 Comme le CO2 est soit le point de départ de la photosynthèse, soit le point final de l’oxydation du substrat, on sait maintenant que les anhydrases carboniques sont omniprésentes et qu’elles sont présentes chez les animaux, les plantes, les bactéries et les champignons. Des enzymes différentes provenant de sources différentes, catalysant la même réaction et ayant généralement des structures homologues, sont appelées isoenzymes. Jusqu’à présent, un total de 7 classes distinctes d’AC a été identifié en fonction de l’organisme : alpha, beta, gamma, delta, zeta, eta et theta. Chaque classe peut contenir plusieurs isoenzymes. Parfois, un même organisme possède plus d’une isoenzyme pour une fonction particulière, comme c’est le cas pour l’anhydrase carbonique humaine. Les humains possèdent 15 CA qui appartiennent à la classe alpha ; ces isoenzymes varient selon leur localisation dans l’organisme et leur activité catalytique. L’AC est un exemple classique d’enzyme hydrolytique, qui catalyse l’ajout ou le retrait d’eau à une molécule de substrat. Plus précisément, l’AC catalyse la conversion réversible du dioxyde de carbone (CO2) en bicarbonate (HCO3-), également appelé acide carbonique.

Bien que l’hydratation du CO2 soit spontanée dans l’eau à pH 7, la réaction est cinétiquement lente (k = 10-1 s-1), trop lente pour convertir tout le CO2 produit par la respiration. Ce n’est qu’au-dessus de pH 9 que la réaction non catalysée devient rapide, en raison de l’attaque directe de OH-, qui est un bien meilleur nucléophile que H2O (k = 104 M-1 s-1, où M-1 se réfère à la concentration de OH-). La figure ci-dessous compare l’attaque nucléophile de l’eau par rapport à celle de l’hydroxyde (OH-) sur le CO2.
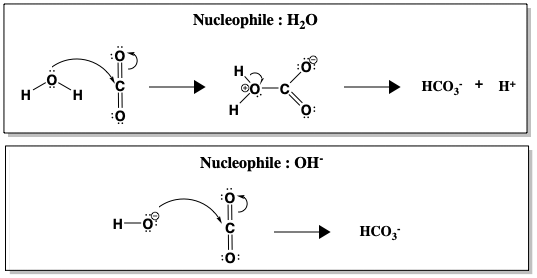
Entre H2O et OH-, la formation de HCO3- est plus rapide lorsque OH- est le nucléophile. Une réaction plus rapide à un pH plus élevé, lorsque plus d’ions OH- sont présents, suggère que OH- est impliqué dans l’étape déterminant la vitesse. Cependant, d’un point de vue réaliste, le pH du sang humain ne peut être modifié pour accélérer l’hydratation du CO2. Au lieu de cela, les humains utilisent l’anhydrase carbonique pour catalyser la réaction. Lorsque l’AC est présente, la réaction est accélérée à un taux de k = 106 s-1.
L’omniprésence de l’AC dans différents organismes reflète l’importance de ces enzymes dans le maintien de la vie. La vitesse d’hydratation du CO2 catalysée par l’AC est essentielle pour répondre aux besoins des cellules vivantes. Parmi les fonctions physiologiques de l’AC figurent la régulation du pH, la sécrétion d’électrolytes, le transport d’ions et l’homéostasie du CO2. Dans le tube digestif, l’AC joue un rôle dans la sécrétion d’acide et maintient la neutralité de la salive en modulant le pH.G,H Parmi ces fonctions, l’AC joue notamment un rôle dans le transport du CO2 et du HCO3- lié à la respiration, le processus d’échange d’oxygène et de dioxyde de carbone atmosphérique qui se produit lorsque les humains inspirent de l’oxygène et expirent du dioxyde de carbone. Peu soluble, le CO2 doit être converti en une forme plus soluble, le HCO3-, pour être transporté dans l’organisme. L’ion bicarbonate (HCO3-) finit par atteindre les poumons, se reconvertit en CO2 et sort du corps par l’expiration.C
La recherche médicale qui tourne autour de l’AC se concentre sur le site actif contenant du Zn comme cible thérapeutique pour le traitement de diverses maladies ; les inhibiteurs et les activateurs de l’AC sont incorporés dans la conception des médicaments. Les inhibiteurs de l’AC sont utilisés comme traitement de l’épilepsie, des ulcères, du cancer, de l’obésité et d’autres troubles neurologiques. Dans l’œil, l’AC produit des ions hydrogène qui maintiennent la pression optique. Cependant, une pression trop élevée dans l’œil peut endommager le nerf optique et provoquer un glaucome. L’activité de l’AC peut créer un gradient de concentration qui entraîne le transport d’eau vers le nerf optique. Lorsqu’il y a trop d’eau autour du nerf optique, la pression autour du nerf augmente et provoque des dommages. L’inhibition de l’AC est devenue un traitement clé du glaucome.
Au-delà des applications pharmaceutiques, le CA a également été étudié pour son utilité dans les applications de capture et de capteur de carbone. La capture et le stockage du carbone se produisent lorsque les CA convertissent le CO2 en bicarbonate. La disponibilité accrue de bicarbonate en présence d’ions calcium (Ca2+) entraîne la précipitation de calcite (CaCO3). Ce processus, appelé bio-minéralisation, pourrait être un mode viable de stockage à long terme du CO2 dans la calcite pour atténuer les émissions de CO2.
Les détails de la structure et de la fonction du site actif de CA contenant du Zn ont été élucidés au cours de 80 années de recherche. L’article actuel approfondit le site actif du métalloenzyme en utilisant des concepts bioinorganiques.
Old Text Below ————————————————————————————————————————————————————————————————–
L’hydratation du dioxyde de carbone et son mécanisme dans les systèmes vivants sont d’une importance fondamentale pour la chimie bioinorganique. En 1932, l’existence d’une enzyme catalysant l’hydratation du CO2 dans les globules rouges a été établie,31 L’enzyme a été nommée anhydrase carbonique (en abrégé CA). En 1939, il a été reconnu que l’enzyme contenait du zinc.32 Comme le CO2 est soit le point de départ de la photosynthèse, soit le point final de l’oxydation du substrat, on sait maintenant que les anhydrases carboniques sont omniprésentes et qu’elles sont présentes chez les animaux, les plantes et plusieurs bactéries. Des enzymes différentes provenant de sources différentes, catalysant la même réaction et ayant généralement des structures homologues, sont appelées isoenzymes. Parfois, un même organisme possède plus d’une isoenzyme pour une fonction particulière, comme c’est le cas pour l’anhydrase carbonique humaine.
Le CO2 gazeux est relativement soluble dans l’eau (3 x 10-2 M à température ambiante sous pCO2 = 1 atm), s’équilibrant avec le carbonate d’hydrogène au pKa 6.1:
\
La réaction non catalysée est cinétiquement lente autour du pH physiologique (k \(\simeq\) 10-1 s-1), alors que, en présence de l’isoenzyme la plus efficace de l’AC, le nombre maximal de rotation du CO2 (c’est-à-dire, le nombre de molécules de substrat transformées par unité de temps par chaque molécule d’enzyme)33 est de \(\simeq\) 106 s-1. L’attaque non catalysée par l’eau sur le CO2 peut être facilitée par deux molécules d’eau liées à l’hydrogène, dont l’une active le carbone au moyen d’une liaison hydrogène avec un oxygène terminal du CO2, et l’autre lie l’atome de carbone via l’oxygène : 34,35
\(\tag{2.7}\)
C’est seulement au-dessus de pH 9 que la réaction non catalysée devient rapide, en raison de l’attaque directe de OH-, qui est un bien meilleur nucléophile que H2O (k \(\simeq\) 104 M-1s-1, où M-1 désigne la concentration de OH-) :
En revanche, la constante de vitesse en présence de l’enzyme, appelée kcat, est indépendante du pH au-delà de pH 8 dans chaque isoenzyme de l’AC (figure 2.2).33,36
In vitro, l’anhydrase carbonique est assez polyvalente, catalysant plusieurs réactions qui impliquent à la fois OH- et H+, comme l’hydrolyse des esters et l’hydratation des aldéhydes. Les diverses isoenzymes ont été caractérisées à différents degrés de sophistication. Les formes à haute activité sont appelées II (kcat \(\simeq\) 106 s-1 à 25 °C) ; les formes à faible activité I (kcat \(\simeq\) 105 s-1), et les formes à très faible activité III (kcat \(\simeq\) 103 s-1).37 Des informations structurelles aux rayons X à une résolution nominale de 2 Å sont disponibles pour l’HCA I38 et l’HCA II,39 où H indique l’homme. La structure de l’HCA II a été affinée récemment.40 Des structures à haute résolution de mutants et de leurs dérivés substrats et inhibiteurs sont rapportées.211 Toutes les isoenzymes sont des polypeptides à chaîne unique, avec un poids moléculaire d’environ 30 kDa et un ion zinc par molécule. Elles ont la forme d’un ballon de rugby avec une crevasse de 16 Å de profondeur traversant le pôle sud (Figure 2.3 Voir la section des plaques en couleur, page C-2.). Au fond de la crevasse, l’ion zinc est ancré à la protéine par trois atomes d’azote d’histidine et est exposé au solvant. Deux histidines (His-94 et His-96, numérotation HCA I) sont liées au zinc par leurs atomes N\(\epsilon\)2, tandis qu’une (His-119) est liée par son atome N\(\delta\)1 (figure 2.4). Il est assez général que les histidines fixent le zinc aussi bien par l’un ou l’autre des deux azotes de l’histidine, la préférence étant probablement dictée par les contraintes stériques imposées par le repliement de la protéine. Les trois protons NH de l’histidine sont tous engagés dans des liaisons H (figure 2.4). L’histidine-119 est impliquée dans une liaison H avec un résidu glutamate. Comme mentionné, cela pourrait être un moyen de contrôler la basicité des ligands métalliques. Une molécule de solvant liée au zinc est impliquée dans une liaison H avec Thr-199, qui à son tour est lié par une liaison H à Glu-106. Ce réseau de liaisons H est important pour comprendre les changements structurels subtils qui se produisent avec les changements de pH ; ceux-ci pourraient, en principe, expliquer les propriétés dépendantes du pH. Bien que la structure des cristaux cultivés à pH 8 dans un tampon contenant du sulfate donne une certaine indication d’une seule molécule de solvant liée au zinc (Figures 2.3 et 2.5 ; voir la section des plaques en couleur, pages C2, C3.), des études théoriques indiquent que deux molécules d’eau peuvent se trouver à des distances de liaison42. Une telle découverte est cohérente avec les études spectroscopiques sur d’autres dérivés et avec le concept selon lequel l’attachement et le détachement des substrats se produisent par le biais de cinq coordinations.
Comme c’est le cas pour chaque enzyme dans laquelle le zinc se trouve au niveau du site catalytique, l’activité est perdue si le métal est retiré, et est restaurée par l’absorption de zinc. La structure tertiaire de l’anhydrase carbonique est maintenue en l’absence de zinc ; même l’apoprotéine dénaturée peut se replier spontanément d’une bobine aléatoire à une conformation native. Bien qu’un tel processus soit accéléré par le zinc,43,44 la présence du métal ne semble pas être une condition absolue pour le repliement correct de l’AC, alors qu’elle l’est pour plusieurs autres métalloprotéines23. ,29,30
Les anions sont attirés dans la cavité métallique par la partie positive Zn(N3OH2)2+, et on pense qu’ils se lient très efficacement au zinc dans l’anhydrase carbonique ; leur utilisation doit donc être évitée autant que possible si le but est d’étudier l’enzyme telle qu’elle est. Lorsque la protéine est dialysée contre de l’eau fraîchement doublement distillée ou soigneusement déionisée sous une atmosphère inerte, le pH de l’échantillon approche le point isoélectrique, qui est inférieur à 6 pour les enzymes HCA I et bovine (BCA II). Le pH peut alors être ajusté par des additions appropriées de NaOH. Toutes les mesures rapportées dans la littérature effectuées dans des tampons d’acétate, de phosphate, d’imidazole ou de trisulfate sont affectées par l’interférence de l’anion avec l’ion métallique. Cependant, les espèces tampons contenant de grands anions comme Hepes (acide 4éthanesulfonique) peuvent être utilisées,45 car ces anions ne pénètrent pas dans la cavité.
Il existe de nombreuses indications que le zinc dans la forme à pH élevé de l’AC est à quatre coordinations avec un groupe OH dans le quatrième site de coordination. À faible pH, l’enzyme existe sous une forme qui contient de l’eau coordonnée ; le nombre de coordination peut être de quatre (une molécule d’eau) ou de cinq (deux molécules d’eau). Bien sûr, l’apparition de l’espèce à faible pH dépend des pKa des équilibres acide-base complexes.
.