はじめに
生体内における二酸化炭素(CO2)の水和とそのメカニズムは、生物無機化学にとって基本的に重要である。 1932年、赤血球中に二酸化炭素の水和を触媒する酵素の存在が確認され31、この酵素はcarbonic anhydrase(CAと略す)と名付けられた。 1939年、この酵素には亜鉛(Zn)が含まれていることが確認された32。CO2は光合成の起点でもあり、基質の酸化の終点でもあることから、炭酸脱水酵素は動物、植物、細菌、真菌に至るまで、どこにでも存在することが知られている。 同じ反応を触媒し、通常は相同な構造を持つ、異なる供給源からの異なる酵素は、アイソザイムと呼ばれる。 これまでのところ、CAは生物によってα、β、γ、δ、Ζ、η、θの7つのクラスに分類されている。 それぞれのクラスには複数のアイソザイムが存在する。 ヒトの炭酸脱水酵素のように、同じ生物が特定の機能に対して2つ以上のアイソザイムを持つこともある。 ヒトには、アルファクラスに属する15種類のCAが存在する。これらのアイソザイムは、体内の場所や触媒活性によって異なる。 CAは加水分解酵素の典型的な例であり、基質分子に水を加えたり取り除いたりする作用を持つ。 より具体的には、CAは二酸化炭素(CO2)を炭酸とも呼ばれる重炭酸(HCO3-)に可逆的に変換することを触媒する。

CO2の水和はpH7の水中では自発的に起こりますが、反応速度は遅く(k = 10-1 s-1)、呼吸で発生したすべてのCO2を変換するには遅すぎます。 pH9以上になると、H2Oよりもはるかに優れた求核剤であるOH-が直接攻撃してくるため、無触媒の反応が高速になる(k = 104 M-1 s-1、M-1はOH-濃度)。 下の図は、CO2に対する水と水酸化物(OH-)の求核性攻撃を比較したものです。
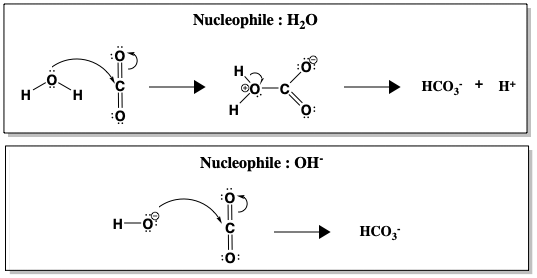
H2OとOH-の間では、OH-を求核剤とした方がHCO3-の生成が速くなります。 pHが高いほど、より多くのOH-イオンが存在するほど反応が速くなることから、OH-が反応速度を決定するステップに関与していると考えられます。 しかし、現実的には、人間の血液のpHを変えてCO2の水和を速めることはできない。 その代わりに、人間は炭酸脱水酵素を使って反応を触媒している。
CAが様々な生物に遍在していることは、この酵素が生命維持に重要であることを示している。
CAが生物界に広く存在することは、この酵素が生命維持に重要な役割を果たしていることを示している。 CAの生理的機能としては、pH調整、電解質分泌、イオン輸送、CO2ホメオスタシスなどが挙げられる。 G,H これらの機能の中でも、CAは、人間が酸素を吸って二酸化炭素を吐くという大気中の酸素と二酸化炭素の交換プロセスである呼吸に関連するCO2とHCO3-の輸送に最も重要な役割を果たしている。 溶解度の低いCO2は、溶解度の高いHCO3-に変換されて全身に運ばれることになる。
CAを中心とした医学研究では、Znを含む活性部位をさまざまな疾患の治療ターゲットとして注目しており、CAの阻害剤と活性化剤の両方がドラッグデザインに組み込まれている。 CAの阻害剤は、てんかん、潰瘍、がん、肥満、その他の神経疾患などの治療に用いられている。 目の中では、CAは水素イオンを生成して視圧を維持している。 しかし、眼球内の圧力が高すぎると、視神経が損傷し、緑内障の原因となる。 CAの活動により、濃度勾配が生じ、水が視神経に運ばれる。 視神経の周りに水が多くなると、視神経の周りの圧力が上昇し、障害が発生します。 CAを阻害することは、緑内障の重要な治療法となっています。
医薬品への応用以外にも、CAは炭素回収や炭素センサーへの応用も検討されています。 炭素回収・貯留は、CAがCO2を重炭酸塩に変換することで起こります。 カルシウムイオン(Ca2+)の存在下で重炭酸塩が増加すると、方解石(CaCO3)が析出する。 このプロセスはバイオミネラル化と呼ばれ、CO2の排出を抑制するためにCO2を方解石に長期的に貯蔵する方法として有効であると考えられる。
CAのZnを含む活性部位の構造と機能の詳細は、80年以上の研究によって明らかにされてきた。
CAのZnを含む活性部位の構造と機能の詳細は、80年以上の研究により解明されていますが、今回は生物無機学的な概念を用いて金属酵素の活性部位を掘り下げてみました。
以下、古い文章 ————————————————————————————————————————————————————————————————–
生体内における二酸化炭素の水和とそのメカニズムは、生物無機化学にとって基本的な重要性を持っています。 1932年、赤血球中に二酸化炭素の水和を触媒する酵素の存在が確認され31、この酵素はcarbonic anhydrase(略称CA)と名付けられた。 1939年、この酵素には亜鉛が含まれていることが確認された32。CO2は光合成の起点でもあり、基質の酸化の終点でもあることから、炭酸脱水酵素は動物、植物、そしていくつかの細菌に至るまで、どこにでも存在することが知られている。 同じ反応を触媒し、通常は相同な構造を持つ、異なる供給源からの異なる酵素は、アイソザイムと呼ばれる。
CO2ガスは水に比較的溶けやすく(室温、pCO2=1気圧で3×10-2M)、炭酸水素塩と平衡してpKa6となります。
無触媒の反応は生理的pH付近では速度が遅く(k˶˶)10-1 s-1ですが、CAの最も効率的なアイソザイムの存在下では、最大CO2ターンオーバー数(=1回のCO2変換で基質分子が変化する数)が大きくなります。 一方、CAの最も効率の良いアイソザイムの存在下では、最大CO2回転数(1分子の酵素によって単位時間当たりに変換される基質分子の数)33は ˶ˆ꒳ˆ˵ ) である。 水によるCO2の無触媒攻撃は、水素結合した2つの水分子によって促進される可能性があります。1つの水分子はCO2の末端酸素と水素結合して炭素を活性化し、もう1つの水分子は酸素を介して炭素原子を結合します。7})
無触媒反応が高速になるのは、pH9以上になってからです。これは、H2Oよりもはるかに優れた求核剤であるOH-の直接攻撃によるものです(k ˶ˆ꒳ˆ˵ 104 M-1s-1、M-1はOH-濃度)。
一方、酵素の存在下での速度定数(kcat)は、どのCAアイソザイムもpH8以上ではpHに依存しません(図2.2).33,36
in vitroでは、炭酸脱水酵素は非常に汎用性が高く、エステルの加水分解やアルデヒドの水和など、OH-とH+の両方が関与するいくつかの反応を触媒している。 様々なアイソザイムは、それぞれ異なる程度に特徴づけられている。 高活性型はII(kcat ˶ˆ꒳ˆ˵)、低活性型はI(kcat ˶ˆ꒳ˆ˵)、超低活性型はIII(kcat ˶ˆ꒳ˆ˵)と呼ばれている37。 211 すべてのアイソザイムは一本鎖のポリペプチドで、M.W.は約30kDa、1分子に1つの亜鉛イオンを持っている。 ラグビーボールのような形をしており、南極には深さ16Åの裂け目がある(図2.3 カラープレートの項、C-2ページ参照)。 すき間の底では、亜鉛イオンは3つのヒスチジン窒素原子によってタンパク質に固定され、溶媒にさらされている。 2つのヒスチジン(His-94とHis-96、HCA Iの番号)はN\\2原子を介して、1つのヒスチジン(His-119)はN\1原子を介して亜鉛と結合している(図2.4)。 一般的に、ヒスチジンは2つのヒスチジンニトロゲンのどちらを使っても同じように亜鉛と結合するが、その優先順位はおそらくタンパク質のフォールディングによって課される立体的な制約によって決定される。 3つのヒスチジンNHプロトンはすべてH結合に関与している(図2.4)。 ヒスチジン-119はグルタミン酸残基とのH結合に関与している。 前述のように、これは金属リガンドの塩基性をコントロールする方法であると考えられる。 亜鉛と結合している溶媒分子は、Thr-199とのH結合に関与しており、さらにそのThr-199はGlu-106とH結合している。 このH結合ネットワークは、pHの変化に伴って生じる微妙な構造変化を理解する上で重要であり、原理的にはpH依存性の特性を説明することができる。 硫酸塩を含む緩衝液を用いてpH8で成長させた結晶の構造は、亜鉛に結合している溶媒分子が1つであることを示唆しているが(図2.3および2.5、カラープレートセクション、ページC2、C3参照)、理論的な研究によると、2つの水分子が結合している可能性があることがわかっている42。
亜鉛が触媒部位にあるすべての亜鉛酵素に当てはまるように、金属が取り除かれると活性が失われ、亜鉛を取り込むことで回復するのである。 炭酸脱水酵素の三次構造は、亜鉛がない状態でも維持され、変性したアポタンパク質でさえ、ランダムなコイルからネイティブに近い構造へと自発的にリフォールドすることができる。 このようなプロセスは亜鉛によって促進されるが43,44、他のいくつかの金属タンパク質では亜鉛の存在が絶対条件であるのに対し、CAの正しいフォールディングには亜鉛の存在は絶対条件ではないようだ23,29,30。 23,29,30
アニオンは金属キャビティ内でポジティブなZn(N3OH2)2+部分に引き寄せられ、炭酸脱水酵素の亜鉛と非常に効果的に結合すると考えられています。そのため、酵素をそのまま研究することが目的であれば、アニオンの使用はできる限り避けるべきです。 タンパク質を不活性雰囲気下で新鮮な二重蒸留水または慎重な脱イオン水に対して透析すると、サンプルのpHは等電点に近づき、HCA Iおよびウシ(BCA II)の酵素では6以下となる。 その後,適切な量のNaOHを加えてpHを調整することができる。 酢酸塩,リン酸塩,イミダゾール,硫酸トリスの各緩衝液で行った文献に記載されている測定値はすべて,金属イオンに対するアニオンの干渉の影響を受けます。
高pH形態のCAの亜鉛は、4番目の配位部位にOH基を持つ4配位であることを示す多くの証拠があります。 低pHでは、酵素は配位した水を含む形で存在し、その配位数は4(水分子1つ)または5(水分子2つ)となります。 もちろん、低pHの種の発生は、複雑な酸塩基平衡のpKaに依存します。