Einführung
Die Kohlendioxid (CO2)-Hydratisierung und ihr Mechanismus in lebenden Systemen sind von fundamentaler Bedeutung für die bioanorganische Chemie. Im Jahr 1932 wurde die Existenz eines Enzyms nachgewiesen, das die CO2-Hydratisierung in roten Blutkörperchen katalysiert,31 Das Enzym wurde Carbonic Anhydrase (abgekürzt CA) genannt. Im Jahr 1939 wurde erkannt, dass das Enzym Zink (Zn) enthält.32 Da CO2 entweder der Ausgangspunkt für die Photosynthese oder der Endpunkt der Substratoxidation ist, weiß man heute, dass Kohlensäureanhydrasen ubiquitär sind und in Tieren, Pflanzen, Bakterien und Pilzen vorkommen. Verschiedene Enzyme aus unterschiedlichen Quellen, die die gleiche Reaktion katalysieren und in der Regel homologe Strukturen aufweisen, werden als Isoenzyme bezeichnet. Bisher wurden insgesamt 7 verschiedene Klassen von CAs basierend auf Organismen identifiziert: alpha, beta, gamma, delta, zeta, eta und theta. Jede Klasse kann mehrere Isoenzyme enthalten. Manchmal hat ein und derselbe Organismus mehr als ein Isoenzym für eine bestimmte Funktion, wie z. B. bei der menschlichen Kohlensäureanhydrase. Der Mensch verfügt über 15 CAs, die zur Alpha-Klasse gehören; diese Isoenzyme unterscheiden sich durch ihre Lage im Körper und durch ihre katalytische Aktivität. CA ist ein klassisches Beispiel für ein hydrolytisches Enzym, eines, das die Addition oder Entfernung von Wasser an ein Substratmolekül katalysiert. Genauer gesagt katalysiert CA die reversible Umwandlung von Kohlendioxid (CO2) in Bicarbonat (HCO3-), auch als Kohlensäure bezeichnet.

Obwohl die Hydratisierung von CO2 in Wasser bei pH 7 spontan abläuft, ist die Reaktion kinetisch langsam (k = 10-1 s-1), zu langsam, um das gesamte bei der Atmung entstehende CO2 umzusetzen. Erst oberhalb von pH 9 wird die unkatalysierte Reaktion schnell, was auf den direkten Angriff von OH- zurückzuführen ist, das ein viel besseres Nukleophil als H2O ist (k = 104 M-1 s-1, wobei sich M-1 auf die OH- Konzentration bezieht). Die folgende Abbildung vergleicht den nukleophilen Angriff von Wasser gegenüber Hydroxid (OH-) an CO2.
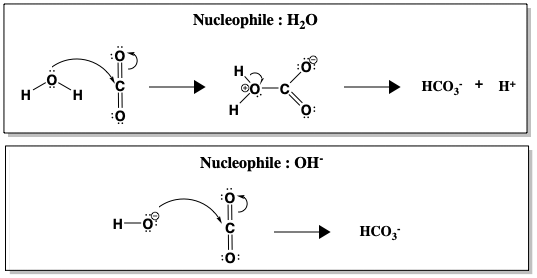
Zwischen H2O und OH- erfolgt die Bildung von HCO3- schneller, wenn OH- das Nukleophil ist. Eine schnellere Reaktion bei höherem pH-Wert, wenn mehr OH–Ionen vorhanden sind, deutet darauf hin, dass OH- an dem geschwindigkeitsbestimmenden Schritt beteiligt ist. Realistischerweise kann der pH-Wert des menschlichen Blutes jedoch nicht verändert werden, um die Hydratation von CO2 zu beschleunigen. Stattdessen katalysiert der Mensch die Reaktion mit Hilfe von Carbonischer Anhydrase. Wenn CA vorhanden ist, wird die Reaktion auf eine Rate von k = 106 s-1 beschleunigt.
Die Ubiquität von CA in verschiedenen Organismen spiegelt die Bedeutung dieser Enzyme für die Aufrechterhaltung des Lebens wider. Die Geschwindigkeit der CA-katalysierten CO2-Hydratisierung ist essentiell, um die Bedürfnisse der lebenden Zellen zu erfüllen. Einige physiologische CA-Funktionen umfassen pH-Regulierung, Elektrolyt-Sekretion, Ionentransport und CO2-Homöostase. Im Verdauungstrakt spielen CAs eine Rolle bei der Sekretion von Säure und halten den Speichel neutral, indem sie den pH-Wert modulieren.G,H Unter diesen Funktionen spielt CA vor allem eine Rolle beim Transport von CO2 und HCO3- im Zusammenhang mit der Atmung, dem Prozess des atmosphärischen Sauerstoff- und Kohlendioxid-Austauschs, der stattfindet, wenn Menschen Sauerstoff einatmen und Kohlendioxid ausatmen. Aufgrund seiner geringen Löslichkeit muss CO2 für den Transport durch den Körper in eine besser lösliche Form, HCO3-, umgewandelt werden. Das Bicarbonat-Ion (HCO3-) erreicht schließlich die Lunge, wird wieder in CO2 umgewandelt und verlässt den Körper durch die Ausatmung.C
Die medizinische Forschung rund um CA konzentriert sich auf die Zn-haltige aktive Stelle als therapeutisches Ziel für verschiedene Krankheitsbehandlungen; sowohl CA-Inhibitoren als auch Aktivatoren werden in das Medikamentendesign einbezogen. CA-Inhibitoren werden zur Behandlung von Epilepsie, Geschwüren, Krebs, Fettleibigkeit und anderen neurologischen Erkrankungen eingesetzt. Im Auge produziert CA Wasserstoffionen, die den Augeninnendruck aufrechterhalten. Ein zu hoher Druck im Auge kann jedoch den Sehnerv schädigen und ein Glaukom verursachen. Die CA-Aktivität kann einen Konzentrationsgradienten erzeugen, der den Transport von Wasser zum Sehnerv antreibt. Wenn sich zu viel Wasser um den Sehnerv herum befindet, erhöht sich der Druck um den Nerv herum, was zu Schäden führt. Die Hemmung von CA hat sich zu einem Schlüssel für die Behandlung des Glaukoms entwickelt.
Neben pharmazeutischen Anwendungen wurde CA auch auf seine Nützlichkeit bei der Kohlenstoffabscheidung und -sensorik untersucht. Die Abscheidung und Speicherung von Kohlenstoff geschieht, indem CAs CO2 in Bikarbonat umwandeln. Die erhöhte Verfügbarkeit von Bicarbonat in Gegenwart von Calcium-Ionen (Ca2+) führt zur Ausfällung von Calcit (CaCO3). Dieser Prozess, der als Bio-Mineralisierung bezeichnet wird, könnte eine praktikable Methode zur langfristigen Speicherung von CO2 in Kalzit sein, um die CO2-Emissionen zu verringern.
Die Details über die Struktur und Funktion der Zn-haltigen aktiven Stelle von CA wurden in 80 Jahren Forschung aufgeklärt. Der aktuelle Artikel beleuchtet das aktive Zentrum des Metalloenzyms mit Hilfe bioanorganischer Konzepte.
Alter Text unter ————————————————————————————————————————————————————————————————–
Die Kohlendioxidhydratation und ihr Mechanismus in lebenden Systemen sind für die bioanorganische Chemie von grundlegender Bedeutung. Im Jahr 1932 wurde die Existenz eines Enzyms nachgewiesen, das die CO2-Hydratisierung in roten Blutkörperchen katalysiert,31 Das Enzym wurde Carbonic Anhydrase (abgekürzt CA) genannt. Im Jahr 1939 wurde erkannt, dass das Enzym Zink enthält.32 Da CO2 entweder der Ausgangspunkt für die Photosynthese oder der Endpunkt der Substratoxidation ist, weiß man heute, dass Kohlensäureanhydrasen ubiquitär sind und in Tieren, Pflanzen und einigen Bakterien vorkommen. Verschiedene Enzyme aus unterschiedlichen Quellen, die dieselbe Reaktion katalysieren und in der Regel homologe Strukturen aufweisen, werden als Isoenzyme bezeichnet. Manchmal besitzt ein und derselbe Organismus mehr als ein Isoenzym für eine bestimmte Funktion, wie z. B. bei der menschlichen Kohlensäureanhydrase.
CO2-Gas ist in Wasser relativ gut löslich (3 x 10-2 M bei Raumtemperatur unter pCO2 = 1 atm), wobei es mit Hydrogencarbonat bei pKa 6 im Gleichgewicht steht.1:
Die unkatalysierte Reaktion ist bei physiologischem pH-Wert kinetisch langsam (k \(\simeq\) 10-1 s-1), während in Gegenwart des effizientesten Isoenzyms von CA die maximale CO2-Umsatzzahl (d.h., die Anzahl der Substratmoleküle, die pro Zeiteinheit von jedem Molekül des Enzyms umgewandelt werden)33 bei \(\simeq\) 106 s-1 liegt. Der unkatalysierte Angriff von Wasser auf CO2 kann durch zwei wasserstoffgebundene Wassermoleküle erleichtert werden, von denen eines den Kohlenstoff über eine Wasserstoffbindung an einen terminalen CO2-Sauerstoff aktiviert, das andere das Kohlenstoffatom über Sauerstoff bindet: 34,35
\(\tag{2.7}\)
Nur oberhalb von pH 9 wird die unkatalysierte Reaktion schnell, aufgrund des direkten Angriffs von OH-, das ein viel besseres Nukleophil als H2O ist (k \(\simeq\) 104 M-1s-1, wobei sich M-1 auf die OH- Konzentration bezieht):
Die Geschwindigkeitskonstante in Gegenwart des Enzyms, kcat genannt, ist dagegen oberhalb von pH 8 bei jedem CA-Isoenzym pH-unabhängig (Abbildung 2.2).33,36
In vitro ist die Carbonische Anhydrase recht vielseitig und katalysiert mehrere Reaktionen, an denen sowohl OH- als auch H+ beteiligt sind, wie z. B. die Hydrolyse von Estern und die Hydratation von Aldehyden. Die verschiedenen Isoenzyme sind unterschiedlich gut charakterisiert worden. Hochaktive Formen werden als II bezeichnet (kcat \(\simeq\) 106 s-1 bei 25 °C), niedrigaktive Formen als I (kcat \(\simeq\) 105 s-1) und die sehr niedrigaktiven Formen als III (kcat \(\simeq\) 103 s-1).37 Röntgenstrukturinformationen mit nominaler 2 Å-Auflösung sind für HCA I38 und HCA II,39 wobei H für Human steht, verfügbar. Die Struktur von HCA II wurde kürzlich verfeinert.40 Hochaufgelöste Strukturen von Mutanten und ihren Substrat- und Inhibitorderivaten werden berichtet.211 Alle Isoenzyme sind einkettige Polypeptide mit einem M.W. von etwa 30 kDa und einem Zinkion pro Molekül. Sie haben die Form eines Rugbyballs mit einem 16 Å tiefen Spalt, der durch den Südpol verläuft (Abbildung 2.3 Siehe Farbtafelabschnitt, Seite C-2.). Am Boden des Spaltes ist das Zink-Ion durch drei Histidin-Stickstoffatome im Protein verankert und dem Lösungsmittel ausgesetzt. Zwei Histidine (His-94 und His-96, HCA I-Nummerierung) sind über ihre N\(\Epsilon\)2-Atome an Zink gebunden, während eines (His-119) über sein N\(\Delta\)1-Atom gebunden ist (Abbildung 2.4). Es ist ziemlich allgemein, dass Histidine Zink gleich gut über jedes der beiden Histidin-Stickstoffatome binden, wobei die Bevorzugung wahrscheinlich durch die sterischen Beschränkungen diktiert wird, die durch die Proteinfaltung auferlegt werden. Die drei Histidin-NH-Protonen sind alle an H-Bindungen beteiligt (Abbildung 2.4). Histidin-119 ist an einer H-Bindung mit einem Glutamat-Rest beteiligt. Wie bereits erwähnt, könnte dies eine Möglichkeit sein, die Basizität der Metallliganden zu kontrollieren. Ein an Zink gebundenes Lösungsmittelmolekül ist in eine H-Bindung mit Thr-199 involviert, das wiederum eine H-Bindung mit Glu-106 hat. Dieses H-Bindungsnetzwerk ist wichtig für das Verständnis der subtilen strukturellen Veränderungen, die bei pH-Änderungen auftreten; diese könnten prinzipiell die pH-abhängigen Eigenschaften erklären. Obwohl die Struktur von Kristallen, die bei pH 8 in sulfathaltigem Puffer gezüchtet wurden, einige Hinweise auf ein einzelnes, an Zink gebundenes Lösungsmittelmolekül gibt (Abbildungen 2.3 und 2.5 Siehe Farbtafelabschnitt, Seiten C2, C3.), deuten theoretische Studien darauf hin, dass sich zwei Wassermoleküle in Bindungsabstand befinden können.42 Ein solcher Befund ist konsistent mit spektroskopischen Untersuchungen an anderen Derivaten und mit dem Konzept, dass die Anlagerung und Ablösung von Substraten durch eine Fünfer-Koordination erfolgt.
Wie bei allen Zinkenzymen, bei denen sich Zink am katalytischen Zentrum befindet, geht die Aktivität verloren, wenn das Metall entfernt wird, und wird durch die Aufnahme von Zink wiederhergestellt. Die Tertiärstruktur der Kohlensäureanhydrase wird in Abwesenheit von Zink beibehalten; sogar das denaturierte Apoprotein kann sich spontan von einer zufälligen Spule zu einer nativen-ähnlichen Konformation zurückfalten. Obwohl ein solcher Prozess durch Zink beschleunigt wird,43,44 scheint die Anwesenheit des Metalls keine absolute Voraussetzung für die korrekte Faltung von CA zu sein, während es für mehrere andere Metalloproteine eine absolute Voraussetzung ist.23 ,29,30
Anionen werden im Metallhohlraum von der positiven Zn(N3OH2)2+-Einheit angezogen und binden vermutlich sehr effektiv an Zink in der Carbonic Anhydrase; daher sollte ihre Verwendung so weit wie möglich vermieden werden, wenn das Ziel darin besteht, das Enzym so zu untersuchen, wie es ist. Wenn das Protein gegen frisch doppelt destilliertes oder sorgfältig deionisiertes Wasser unter inerter Atmosphäre dialysiert wird, nähert sich der pH-Wert der Probe dem isoelektrischen Punkt, der bei HCA I und bovinen (BCA II) Enzymen unter 6 liegt. Der pH-Wert kann dann durch entsprechende Zugabe von NaOH eingestellt werden. Alle in der Literatur berichteten Messungen, die in Acetat-, Phosphat-, Imidazol- oder Tris-Sulfat-Puffern durchgeführt wurden, werden durch die Interferenz des Anions mit dem Metallion beeinflusst. Pufferspezies, die große Anionen wie Hepes (4-Ethansulfonsäure) enthalten, können jedoch verwendet werden,45 da diese Anionen nicht in den Hohlraum eindringen.
Es gibt viele Hinweise darauf, dass Zink in der Hoch-pH-Form von CA vierkoordiniert ist, mit einer OH-Gruppe an der vierten Koordinationsstelle. Bei niedrigem pH-Wert liegt das Enzym in einer Form vor, die koordiniertes Wasser enthält; die Koordinationszahl kann vier (ein Wassermolekül) oder fünf (zwei Wassermoleküle) betragen. Natürlich hängt das Auftreten der Niedrig-pH-Spezies von den pKa’s der komplexen Säure-Basen-Gleichgewichte ab.