Introducción
La hidratación del dióxido de carbono (CO2) y su mecanismo en los sistemas vivos son de importancia fundamental para la química bioinorgánica. En 1932 se estableció la existencia de una enzima que catalizaba la hidratación del CO2 en los glóbulos rojos,31 La enzima se denominó anhidrasa carbónica (abreviada CA). En 1939 se reconoció que la enzima contenía zinc (Zn).32 Dado que el CO2 es el punto de partida de la fotosíntesis o el punto final de la oxidación de los sustratos, ahora se sabe que las anhidrasas carbónicas son ubicuas y se encuentran en animales, plantas, bacterias y hongos. Se denominan isoenzimas a las enzimas de distintas fuentes que catalizan la misma reacción y que suelen tener estructuras homólogas. Hasta ahora, se han identificado un total de 7 clases distintas de AC basadas en el organismo: alfa, beta, gamma, delta, zeta, eta y theta. Cada clase puede contener múltiples isoenzimas. A veces el mismo organismo tiene más de una isoenzima para una función concreta, como ocurre con la anhidrasa carbónica humana. Los seres humanos tienen 15 AC que pertenecen a la clase alfa; estas isozimas varían según la ubicación en el cuerpo y la actividad catalítica. La AC es un ejemplo clásico de enzima hidrolítica, que cataliza la adición o eliminación de agua a una molécula de sustrato. Más concretamente, la AC cataliza la conversión reversible de dióxido de carbono (CO2) en bicarbonato (HCO3-), también denominado ácido carbónico.

Aunque la hidratación del CO2 es espontánea en agua a pH 7, la reacción es cinéticamente lenta (k = 10-1 s-1), demasiado lenta para convertir todo el CO2 producido en la respiración. Sólo por encima de pH 9 la reacción no catalizada se vuelve rápida, debido al ataque directo del OH-, que es un nucleófilo mucho mejor que el H2O (k = 104 M-1 s-1, donde M-1 se refiere a la concentración de OH-). La figura siguiente compara el ataque nucleófilo del agua frente al hidróxido (OH-) sobre el CO2.
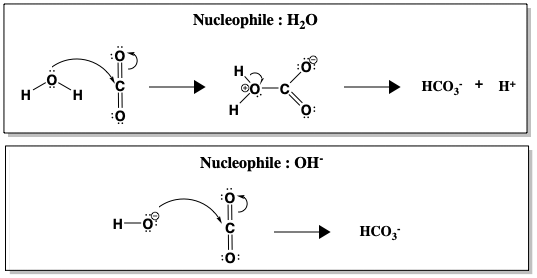
Entre el H2O y el OH-, la formación de HCO3- se produce más rápidamente cuando el OH- es el nucleófilo. Una reacción más rápida a un pH más alto, cuando hay más iones OH- presentes, sugiere que el OH- está involucrado en el paso determinante de la velocidad. Sin embargo, siendo realistas, el pH de la sangre humana no puede modificarse para acelerar la hidratación del CO2. En su lugar, los humanos utilizan la anhidrasa carbónica para catalizar la reacción. Cuando la AC está presente, la reacción se acelera a una velocidad de k = 106 s-1.
La ubicuidad de la AC en diferentes organismos refleja la importancia de estas enzimas para mantener la vida. La velocidad de hidratación del CO2 catalizada por la AC es esencial para satisfacer las necesidades de las células vivas. Algunas funciones fisiológicas de las AC incluyen la regulación del pH, la secreción de electrolitos, el transporte de iones y la homeostasis del CO2. En el tracto digestivo, las AC desempeñan un papel en la secreción de ácido y mantienen la saliva neutra modulando el pH.G,H Entre estas funciones, las AC desempeñan sobre todo un papel en el transporte de CO2 y HCO3- relacionado con la respiración, el proceso de intercambio de oxígeno y dióxido de carbono atmosférico que se produce cuando los seres humanos inhalan oxígeno y exhalan dióxido de carbono. Con una baja solubilidad, el CO2 debe convertirse en una forma más soluble, el HCO3-, para su transporte por todo el cuerpo. El ion bicarbonato (HCO3-) llega finalmente a los pulmones, se convierte de nuevo en CO2 y sale del cuerpo a través de la exhalación.C
La investigación médica que gira en torno al CA se centra en el sitio activo que contiene Zn como objetivo terapéutico para el tratamiento de diversas enfermedades; tanto los inhibidores como los activadores del CA se incorporan al diseño de fármacos. Los inhibidores del CA se utilizan como tratamiento para la epilepsia, las úlceras, el cáncer, la obesidad y otros trastornos neurológicos. En el ojo, la AC produce iones de hidrógeno que mantienen la presión óptica. Sin embargo, un exceso de presión en el ojo puede dañar el nervio óptico y causar glaucoma. La actividad de la AC puede crear un gradiente de concentración que impulsa el transporte de agua hacia el nervio óptico. Cuando hay demasiada agua alrededor del nervio óptico, la presión alrededor del nervio aumenta causando daños. La inhibición de la AC se ha convertido en un tratamiento clave del glaucoma.
Más allá de las aplicaciones farmacéuticas, el CA también se ha investigado por su utilidad en la captura de carbono y en aplicaciones de sensores de carbono. La captura y el almacenamiento de carbono se producen cuando los CA convierten el CO2 en bicarbonato. La mayor disponibilidad de bicarbonato en presencia de iones de calcio (Ca2+) provoca la precipitación de calcita (CaCO3). Este proceso, llamado bio-mineralización, puede ser un modo viable de almacenamiento a largo plazo de CO2 en la calcita para mitigar las emisiones de CO2.
Los detalles sobre la estructura y la función del sitio activo del CA que contiene Zn se han dilucidado a lo largo de 80 años de investigación. El presente artículo profundiza en el sitio activo de la metaloenzima utilizando conceptos bioinorgánicos.
Texto antiguo debajo de ————————————————————————————————————————————————————————————————–
La hidratación del dióxido de carbono y su mecanismo en los sistemas vivos son de importancia fundamental para la química bioinorgánica. En 1932 se estableció la existencia de una enzima que cataliza la hidratación del CO2 en los glóbulos rojos,31 La enzima fue denominada anhidrasa carbónica (abreviada CA). En 1939 se reconoció que la enzima contenía zinc.32 Dado que el CO2 es el punto de partida de la fotosíntesis o el punto final de la oxidación de sustratos, se sabe que las anhidrasas carbónicas son ahora ubicuas y se encuentran en animales, plantas y varias bacterias. Se denominan isoenzimas a las enzimas de diferentes fuentes que catalizan la misma reacción y que suelen tener estructuras homólogas. A veces el mismo organismo tiene más de una isoenzima para una función particular, como es el caso de la anhidrasa carbónica humana.
El gas CO2 es relativamente soluble en agua (3 x 10-2 M a temperatura ambiente bajo pCO2 = 1 atm), equilibrándose con el carbonato de hidrógeno a pKa 6.1:
La reacción no catalizada es cinéticamente lenta alrededor del pH fisiológico (k \(\simeq\) 10-1 s-1), mientras que, en presencia de la isoenzima más eficiente de CA, el número máximo de recambio de CO2 (es decir, el número de moléculas de sustrato transformadas por unidad de tiempo por cada molécula de enzima)33 es \(\simeq\) 106 s-1. El ataque no catalizado del agua sobre el CO2 puede ser facilitado por dos moléculas de agua con enlace de hidrógeno, una de las cuales activa el carbono mediante un enlace de hidrógeno a un oxígeno terminal del CO2, y la otra se une al átomo de carbono a través del oxígeno: 34,35
(\tag{2.7})
Sólo por encima de pH 9 la reacción no catalizada se vuelve rápida, debido al ataque directo del OH-, que es un nucleófilo mucho mejor que el H2O (k \(\simeq\) 104 M-1s-1, donde M-1 se refiere a la concentración de OH-):
Por otro lado, la constante de velocidad en presencia de la enzima, denominada kcat, es independiente del pH por encima de pH 8 en cada isoenzima de CA (Figura 2.2).33,36
In vitro, la anhidrasa carbónica es bastante versátil, catalizando varias reacciones que implican tanto OH- como H+, como la hidrólisis de ésteres y la hidratación de aldehídos. Las distintas isoenzimas se han caracterizado con diferentes grados de sofisticación. Las formas de alta actividad se denominan II (kcat \(\simeq\) 106 s-1 a 25 °C); las formas de baja actividad I (kcat \(\simeq\) 105 s-1), y las formas de muy baja actividad III (kcat \(\simeq\) 103 s-1).37 Se dispone de información estructural de rayos X a una resolución nominal de 2 Å para la HCA I38 y la HCA II,39 donde H indica humano. La estructura de la HCA II ha sido refinada recientemente.40 Se están publicando estructuras de alta resolución de mutantes y de sus derivados de sustrato e inhibidores.211 Todas las isoenzimas son polipéptidos de cadena simple, con un peso molecular de unos 30 kDa y un ion de zinc por molécula. Tienen la forma de un balón de rugby con una hendidura de 16 Å de profundidad que atraviesa el polo sur (Figura 2.3 Véase la sección de láminas en color, página C-2.). En la parte inferior de la hendidura, el ion de zinc está anclado a la proteína por tres átomos de nitrógeno de histidina y está expuesto al disolvente. Dos histidinas (His-94 e His-96, numeración HCA I) están unidas al zinc a través de sus átomos N\(\epsilon\)2, mientras que una (His-119) está unida a través de su átomo N\(\delta\)1 (Figura 2.4). Es bastante general que las histidinas se unan al zinc igualmente bien por cualquiera de los dos nitrógenos de histidina, siendo la preferencia probablemente dictada por las restricciones estéricas impuestas por el plegamiento de la proteína. Los tres protones NH de la histidina participan en el enlace H (Figura 2.4). La histidina-119 participa en el enlace H con un residuo de glutamato. Como se ha mencionado, esto podría ser una forma de controlar la basicidad de los ligandos metálicos. Una molécula de disolvente unida al zinc está implicada en un enlace H con Thr-199, que a su vez está unido por H a Glu-106. Esta red de enlaces H es importante para entender los sutiles cambios estructurales que se producen con los cambios de pH; éstos podrían, en principio, explicar las propiedades dependientes del pH. Aunque la estructura de los cristales cultivados a pH 8 en un tampón que contiene sulfato da algunos indicios de una única molécula de disolvente unida al zinc (Figuras 2.3 y 2.5, véase la sección de láminas en color, páginas C2, C3.), los estudios teóricos indican que dos moléculas de agua pueden estar a distancias de enlace.42 Tal hallazgo es consistente con los estudios espectroscópicos en otros derivados y con el concepto de que la unión y el desprendimiento de los sustratos ocurren a través de cinco coordinaciones.
Al igual que ocurre con todas las enzimas de zinc en las que el zinc se encuentra en el sitio catalítico, la actividad se pierde si se elimina el metal, y se restablece mediante la absorción de zinc. La estructura terciaria de la anhidrasa carbónica se mantiene en ausencia de zinc; incluso la apoproteína desnaturalizada puede replegarse espontáneamente desde una bobina aleatoria hasta una conformación similar a la nativa. Aunque dicho proceso se acelera con el zinc,43,44 la presencia del metal no parece ser un requisito absoluto para el correcto plegamiento de la AC, mientras que sí lo es para otras metaloproteínas.23 ,29,30
Los aniones son atraídos en la cavidad del metal por la fracción positiva Zn(N3OH2)2+, y se cree que se unen al zinc en la anhidrasa carbónica de forma muy efectiva; por lo que su uso debe evitarse en la medida de lo posible si el objetivo es estudiar la enzima tal cual. Cuando la proteína se dializa contra agua recién bidestilada o cuidadosamente desionizada bajo una atmósfera inerte, el pH de la muestra se aproxima al punto isoeléctrico, que es inferior a 6 para las enzimas HCA I y bovina (BCA II). El pH puede entonces ajustarse mediante adiciones adecuadas de NaOH. Todas las mediciones reportadas en la literatura realizadas en tampones de acetato, fosfato, imidazol o trisulfato se ven afectadas por la interferencia del anión con el ion metálico. Sin embargo, pueden utilizarse especies tampón que contengan aniones grandes como Hepes (ácido 4etanosulfónico),45 ya que estos aniones no entran en la cavidad.
Hay muchos indicios de que el zinc en la forma de alto pH de la AC está cuatricordinado con un grupo OH en el cuarto sitio de coordinación. A pH bajo, la enzima existe en una forma que contiene agua coordinada; el número de coordinación puede ser cuatro (una molécula de agua) o cinco (dos moléculas de agua). Por supuesto, la aparición de las especies de bajo pH depende de los pKa de los complejos equilibrios ácido-base.